
13.6: Diferencias de sexo e interpretaciones evolutivas
- Page ID
- 148263
- Explicar las diferencias de sexo que se han reportado en el cerebro, los sistemas sensoriales y las funciones cognitivas, y las limitaciones de estos hallazgos
- Describir cómo las hormonas pueden influir en las diferencias sexuales en el comportamiento, como las preferencias de juguetes infantiles y las conductas agresivas
- Identificar las premisas centrales de la teoría de estrategias sexuales
Visión general
Después de señalar que la dicotomía no refleja realmente las variaciones observadas tanto en el sexo biológico como en la identidad de género, en esta sección se analizan las diferencias de sexo que se han reportado entre hombres y mujeres humanos. Se cubren las diferencias de sexo en el cerebro, las diferencias de sexo en los sistemas sensoriales y cognición, y las diferencias de sexo en el comportamiento, con el último tema que incluye una revisión de las influencias hormonales sobre las diferencias de sexo en el comportamiento seguido de discusiones sobre las preferencias de juguetes infantiles y comportamientos agresivos. Finalmente, se abordan las interpretaciones evolutivas del comportamiento reproductivo, incluyendo la teoría de estrategias sexuales y la teoría del manejo de errores (en lo que se refiere a las conductas reproductivas).
Diferencias de sexo
Nota Preliminar
Secciones anteriores han enfatizado la falta de una dicotomía estricta entre hombres y mujeres, tanto en términos de sexo biológico como de identidad de género. Sin embargo, la mayoría de las investigaciones divide a las personas en dos categorías, más comúnmente basadas en su sexo biológico asignado al nacer, y a menudo también asume que el sexo biológico reportado se correlaciona con la identidad de género. Si bien esto es útil hasta cierto punto, ¡ojalá que la investigación futura tome un enfoque más matizado de los tres aspectos del auto sexo, género y orientación sexual “sexual” de un individuo! A continuación notarás que cuando comparamos a hombres y mujeres en cuanto a rasgos y características psicológicas, en realidad solo hay una ligera diferencia (típicamente no superior al 5%) entre los dos, y solo a nivel grupal. Piénselo de esta manera: los hombres adultos en Estados Unidos son aproximadamente 9% más altos que las mujeres adultas, en promedio (McDowell et al., 2008), y aproximadamente 15% más grandes en masa corporal (Larsen, 2003). Sin embargo, todos pueden pensar en ejemplos de hombres que son más bajos y/o más pequeños que muchas mujeres, y mujeres que son más altas y/o más grandes que muchos hombres- ¡y esta diferencia física es al menos el doble del tamaño de cualquiera de las diferencias que se encuentran en los dominios psicológicos!
Además, como aprenderás en la sección sobre diferencias de sexo en el cerebro, si bien se han reportado diferencias entre cerebros masculinos y femeninos a nivel de grupo, no hay forma de clasificar de manera confiable un cerebro individual como masculino o femenino, debido tanto a la gran superposición entre machos y hembras y al hecho de que cada cerebro tiene su propio mosaico único de rasgos tanto “masculinos” como “femeninos” de maneras diferentes (e impredecibles).
Introducción a las diferencias de sexo
Las gallinas y gallos son diferentes (Figura\(\PageIndex{1}\)). Las vacas y los toros son diferentes. Hombres y mujeres son diferentes. Incluso las niñas y los niños son diferentes. Los humanos, como muchos animales, son sexualmente dimórficos (di, “dos”; morph, “type”) en el tamaño y la forma de sus cuerpos, su fisiología, y para nuestros propósitos, su comportamiento.

Curiosamente, las diferencias físicas en tamaño y apariencia (para muchos individuos, podemos determinar fácilmente el sexo biológico a partir de rasgos visibles) entre hombres y mujeres humanos están relacionadas con algunas de las diferencias que vemos en los comportamientos reproductivos humanos. En el reino animal, muchos animales cuya estrategia de apareamiento es formar parejas monógamas tienen machos y hembras prácticamente indistinguibles, mientras que las especies polígamas (donde un macho suele aparearse con muchas hembras, y algunos machos pierden completamente el apareamiento) son notoriamente diferentes, a menudo tanto en características y en tamaño. Si bien es evidente en algunas especies de aves, como el obviamente diferente gallo y gallina polígamos mencionados anteriormente (versus cisnes monógamos, con machos y hembras casi idénticos), estas diferencias son particularmente evidentes en especies de primates (no humanos) (y muchos otros mamíferos). Ejemplos en especies de primates incluyen los gibones monógamos de apariencia similar (Jaffe, 2019, página 210-211), versus los gorilas polígamos (Jaffe, 2019, página 209), donde los machos adultos son mucho más grandes que las hembras y obtienen la coloración característica de “dorso plateado” a medida que maduran (Figura\(\PageIndex{2}\)). Una diferencia de alrededor del diez por ciento -precisamente donde la especie humana cae en este continuo de diferencia de tamaño- es la línea divisoria entre “monógamo” y “polígamo”; es decir, las especies que tienen menos del diez por ciento de diferencia en apariencia física y tamaño entre machos y hembras suelen ser monógamas y especies que tienen una diferencia mayor al diez por ciento tienden a ser polígamas. Para los humanos, “... los sexos difieren más en los seres humanos que en los mamíferos monógamos, pero mucho menos que en los mamíferos extremadamente polígamos...” (Daly & Wilson, 1996, página 13). Así, no es realmente sorprendente, desde un punto de vista evolutivo, que algunos humanos adopten una estrategia monógama para el apareamiento y la reproducción, y otros sigan una estrategia polígama.
_(2854166549).jpg)
.jpg)
Volviendo al comportamiento humano, el comportamiento de niños y niñas difiere en muchos sentidos. Las niñas generalmente sobresalen en habilidades verbales en relación con los niños; los niños tienen casi el doble de probabilidades que las niñas de sufrir dislexia (dificultades de lectura) y tartamudeo, y casi cuatro veces más probabilidades de sufrir autismo. Los niños son generalmente mejores que las niñas en tareas que requieren habilidades visuoespaciales. Las niñas se involucran en comportamientos de crianza con más frecuencia que los niños. Más del 90% de todos los casos de anorexia nerviosa involucran a mujeres jóvenes. Los hombres jóvenes tienen el doble de probabilidades que las mujeres jóvenes de sufrir esquizofrenia. Los niños son mucho más agresivos físicamente y generalmente participan en juegos más rudos que las niñas (Berenbaum, Martin, Hanish, Briggs, & Fabe, 2008). Muchas diferencias de sexo conductual, como la diferencia en la agresividad física, persisten a lo largo de la edad adulta. Por ejemplo, hay muchos más hombres que mujeres que cumplen penas de prisión por comportamiento violento. A continuación, esta sección aborda las diferencias de sexo en el cerebro, en los sistemas sensoriales y cognición, y en el comportamiento, donde se discuten con mayor detalle las diferencias de sexo en las preferencias de juguete y el comportamiento agresivo mencionado anteriormente.
Diferencias sexuales en el cerebro
Las diferencias de sexo en el tamaño general del cerebro humano se han reportado desde hace años (aunque la verdadera significación de este hallazgo es discutible). Más recientemente, se han descubierto diferencias de sexo en estructuras cerebrales específicas. Por ejemplo, en ratas, el núcleo sexualmente dimórfico (SDN-POA) del área preóptica del hipotálamo es mucho más grande en machos que en hembras. Esto es el resultado de los efectos organizativos de las hormonas esteroides gonadales sobre el cerebro y el comportamiento, que está relativamente limitado a las primeras etapas de desarrollo. En ratas, la exposición a la testosterona (que se convierte en estradiol) o estradiol provoca la masculinización del cerebro. (El mismo mecanismo de una hormona “femenina” masculinizante del cerebro fetal no está presente en los primates, y por lo tanto presumiblemente no en los humanos. La alfa-fetoproteína se une al estradiol en ratas hembra para evitar la masculinización de sus cerebros). La figura\(\PageIndex{3}\) representa cortes transversales a través de los cerebros de ratas que muestran un macho (A, izquierda), una hembra (B, centro) y una hembra tratada con testosterona cuando era recién nacido (C, derecha). Nótese que los SDN-POA (los cuerpos celulares oscuros) del macho son sustancialmente mayores que los de la hembra no tratada pero son iguales en tamaño a los de la hembra tratada con testosterona. El grado en que estas diferencias de sexo en la estructura cerebral explican las diferencias de sexo en el comportamiento permanece sin especificar en los mamíferos.
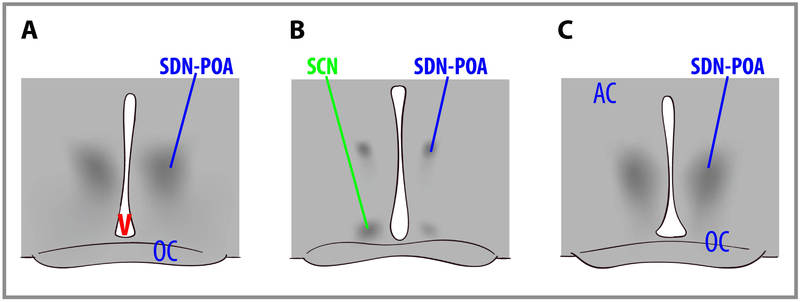
En un área similar al SDN-POA de rata, Simon LeVay (1991) reportó una diferencia comparable en el hipotálamo humano-INAH-3, el tercer núcleo intersticial del hipotálamo anterior. Específicamente, los hombres heterosexuales tenían núcleos INAH-3 que eran más del doble de grandes (por volumen) que los hombres homosexuales, cuyos volúmenes de núcleos INAH-3 eran similares a los de las mujeres. Si bien LeVay pudo replicar el resultado, los intentos de otro investigador solo conducen a una diferencia de nivel de tendencia entre hombres heterosexuales y homosexuales en el volumen del INAH-3 (Byne 2000 & 2001, según lo citado por Bailey et al., 2016). Independientemente del verdadero estado de las posibles diferencias en el núcleo INAH-3 del macho humano (podría ser intermedio entre los hallazgos de los dos investigadores, por ejemplo), “sería poco probable que el tamaño del INAH-3 fuera un factor clave que regula la orientación sexual. Esto se debe a que habría demasiadas excepciones —hombres homosexuales con un INAH-3 grande y hombres heterosexuales con un INAH-3 pequeño— para creer que la talla del INAH-3 es crucial” (Bailey et al., 2016, página 72). El INAH-3 es una estructura diminuta -aproximadamente del tamaño de un grano de arena- y estudiarlo requiere tejido post mortem. Como tal, es poco probable que se estudie más en un futuro próximo (Bailey et al., 2016).
Para características cerebrales más grandes, que pueden ser estudiadas en personas vivas mediante técnicas de imagen, se han reportado muchas diferencias cerebrales entre hombres y mujeres adultos, tanto en regiones corticales como en estructuras subcorticales. Un ejemplo de esto incluye datos de imágenes por resonancia magnética (MRI) que sugieren que las mujeres tienen mayores volúmenes de cortezas paralimbianas frontales y mediales y los hombres tienen mayores volúmenes en la corteza frontomedial, amígdala e hipotálamo (Goldstein et al., 2001). Sin embargo, un escrutinio más detenido a veces hace que estas diferencias sexuales no sean importantes Por ejemplo, se creía ampliamente que el cuerpo calloso difería entre hombres y mujeres, se promocionó que el supuesto aumento de tamaño en las mujeres resultaría en una mayor conectividad entre los hemisferios derecho e izquierdo del cerebro, pero se ha encontrado que no difiere en absoluto cuando hombres y mujeres fueron emparejados para el tamaño del cerebro ( Luders et al., 2014). Otro estudio que subrayó la importancia de tomar en consideración el tamaño del cerebro fue un estudio de gemelos que encontró que una correlación débil entre las diferencias de sexo en el cerebro y las diferencias de sexo conductual resultó ser impulsada principalmente por el tamaño del cerebro. Los autores advirtieron que, por lo tanto, se debe tener precaución al inferir la causalidad (van Eijk et al., 2021).
Adicionalmente, Joel et al. (2015) realizaron un estudio extenso de más de 1400 resonancias magnéticas de cerebros humanos y encontraron que al tomar en cuenta todo el cerebro (y no solo mirar un área específica versus otra, donde se han reportado diferencias de sexo a nivel grupal), es imposible categorizar los cerebros en formas masculinas y femeninas. En cambio, hubo una amplia superposición de características “masculinas” y “femeninas” en los cerebros humanos y era muy raro tener consistencia interna (solo rasgos “masculinos” o solo “femeninos”) en un cerebro específico. Por lo tanto, si bien a menudo podemos categorizar el sexo biológico observando las estructuras anatómicas- a menudo hay diferencias distintas entre los cuerpos masculino y femenino, y un cuerpo dado generalmente tiene solo estructuras masculinas o solo femeninas- no podemos determinar si un cerebro específico vino de un cuerpo masculino o femenino, ni podemos predecir al conocer el sexo/género de un individuo en particular qué forma ha tomado su cerebro. Los autores afirman que “cada cerebro es un mosaico único de características, algunas de las cuales pueden ser más comunes en las hembras en comparación con los machos, otras pueden ser más comunes en los machos en comparación con las hembras, y otras pueden ser comunes tanto en machos como en hembras” (Joel et al., 2015, página 15472). Adicionalmente, sus hallazgos apoyan la noción de que la masculinización y feminización de las áreas cerebrales son dos procesos separados que progresan de manera independiente, permitiendo que las variaciones de diferenciación en un patrón “masculino” o “femenino” ocurran dentro del mismo cerebro.
Diferencias sexuales en sistemas sensoriales y cognición
También se han reportado diferencias de sexo en una serie de funciones sensoriales y cognitivas. Las hembras son generalmente más sensibles a la información auditiva, mientras que los machos son más sensibles a la información visual. Las hembras también suelen ser más sensibles que los machos al gusto y al aporte olfativo. Las mujeres muestran menos lateralización de las funciones cognitivas que los hombres. En promedio, las hembras generalmente sobresalen en habilidades verbales, perceptuales y motrices finas, mientras que los machos superan a las hembras en tareas cuantitativas y visuoespaciales, incluyendo lectura de mapas y búsqueda de dirección. Curiosamente, las mujeres con síndrome de Turner (cuyos ovarios no son funcionales) a menudo tienen memoria espacial deteriorada.
Aunque se pueden documentar diferencias de sexo confiables, estas diferencias en la capacidad son leves. Es importante señalar que hay más variación dentro de cada sexo que entre los sexos para la mayoría de las habilidades cognitivas (Figura\(\PageIndex{4}\)).
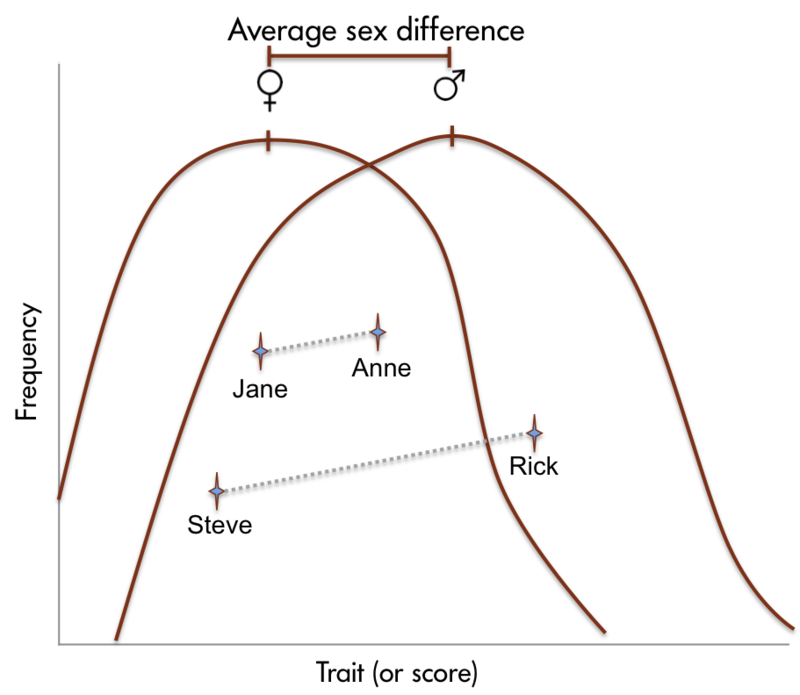
Para enfatizar la importancia de reconocer la vasta variación de rasgos de personalidad y expresiones conductuales que se encuentran dentro de todos los humanos, independientemente de su sexo/género, considere los siguientes resultados del estudio. Joel et al. (2015) realizaron un análisis de los rasgos de personalidad, actitudes, intereses y comportamientos de más de 5,500 individuos. Sus hallazgos reflejaron sus hallazgos de resonancia magnética (gran superposición entre machos y hembras y poca consistencia interna dentro del mismo individuo). Reportan: “De acuerdo con los datos cerebrales, nuestros análisis de datos relacionados con el género revelaron una extensa superposición entre mujeres y hombres en rasgos de personalidad, actitudes, intereses y comportamientos. Además, encontramos que la variabilidad sustancial de las características de género es altamente prevalente, mientras que la consistencia interna es extremadamente rara, incluso para actividades altamente estereotipadas de género... Así, la mayoría de los humanos poseen un mosaico de rasgos de personalidad, actitudes, intereses y comportamientos, algunos más comunes en los machos en comparación con las hembras, otros más comunes en las hembras en comparación con los machos, y aún otros comunes tanto en machos como en hembras” (Joel et al., 2015, página 15472). Es razonable concluir que en general, los hombres y mujeres humanos son más similares de lo que son diferentes, tanto en lo que respecta a las características cerebrales como a los rasgos psicológicos.
Diferencias sexuales en el comportamiento
Revisión de Influencias hormonales en las diferencias sexuales en el comportamiento
Las diferencias hormonales entre hombres y mujeres pueden explicar las diferencias de sexo adulto que se desarrollan durante la pubertad, pero ¿qué explica las diferencias de sexo conductual entre los niños antes de la pubertad y la activación de sus gónadas? Como se discute en la sección 13.2, la presencia (o ausencia) de andrógenos determina si el feto desarrolla una forma corporal masculina o femenina. Los andrógenos de los testículos fetales dirigen el desarrollo del cuerpo, el sistema nervioso central y el comportamiento posterior en dirección masculina. En contraste, los ovarios fetales no secretan altas concentraciones de hormonas, por lo que el cuerpo, el sistema nervioso central, y posteriormente, el comportamiento, siguen una vía femenina.
Como se discute en la sección 13.1, las hormonas esteroides gonadales tienen efectos organizativos (o de programación) sobre el cerebro y el comportamiento (Phoenix, Goy, Gerall, & Young, 1959). Estos efectos organizativos están relativamente limitados a las primeras etapas de desarrollo. En ratas, el tratamiento temprano con hormonas esteroideas provoca una masculinización relativamente irreversible y permanente del comportamiento adulto posterior (apareamiento y agresivo). En contraste, los efectos activacionales de las hormonas (proporcionadas en la edad adulta para influir en los comportamientos) son temporales y reversibles. Así, el comportamiento masculino típico requiere la exposición a andrógenos durante la gestación (en humanos) o inmediatamente después del nacimiento (en ratas) para masculinizar algo el cerebro y también requiere andrógenos durante o después de la pubertad para activar estos circuitos neuronales. El comportamiento típico femenino requiere una falta de exposición a los andrógenos temprano en la vida lo que conduce a la feminización del cerebro y también requiere de estrógenos para activar estos circuitos neuronales en la edad adulta. Pero esta simple dicotomía, que funciona bien con animales con dimorfismo sexual muy distinto en el comportamiento, tiene muchas advertencias cuando se aplica a las personas.
Preferencias de juguetes infantiles
Si caminas por alguna juguetería importante, entonces probablemente observarás un par de pasillos llenos de cajas rosadas y la ausencia total de empaques rosados de juguetes en pasillos adyacentes. Notablemente, también verás una fuerte autosegregación de niños y niñas en estos pasillos. Es raro ver chicos en los pasillos “rosados” y viceversa. A los fabricantes de juguetes a menudo se les acusa de fabricar juguetes que tienen prejuicios de género, pero parece más probable que niños y niñas disfruten jugando con tipos y colores específicos de juguetes. En efecto, los fabricantes de juguetes duplicarían inmediatamente sus ventas si pudieran vender juguetes a ambos sexos. Los niños generalmente prefieren juguetes como camiones y pelotas y las niñas generalmente prefieren juguetes como muñecas. Si bien es dudoso que existan genes que codifiquen preferencias para los coches de juguete y camiones en el cromosoma Y, es posible que las hormonas puedan dar forma al desarrollo del cerebro de un niño para preferir ciertos tipos de juguetes o estilos de comportamiento de juego. También es razonable creer que los niños aprenden qué tipos de juguetes y qué estilos de juego son apropiados a su género (Figura\(\PageIndex{5}\)). ¿Cómo podemos separar la contribución del aprendizaje de los mecanismos fisiológicos subyacentes para comprender las diferencias de sexo en los comportamientos humanos? Para desenredar estos temas, a menudo se utilizan modelos animales. A diferencia de la situación en los humanos, donde las diferencias de sexo suelen ser solo una cuestión de grado (a menudo leve), en algunos animales, los miembros de un solo sexo pueden mostrar un comportamiento particular. Por ejemplo, muchas veces solo cantan los pájaros cantores machos. Los estudios de tales comportamientos fuertemente sesgados por el sexo son particularmente valiosos para comprender la interacción entre el comportamiento, las hormonas y el sistema nervioso.


Un estudio de monos vervet pone en duda la primacía del aprendizaje en el establecimiento de preferencias de juguete (Alexander & Hines, 2002). Los monos vervet hembra preferían los juguetes típicos de las niñas, como muñecas o ollas para cocinar, mientras que los monos vervet machos preferían los juguetes típicos de los niños, como autos o pelotas. No hubo diferencias de sexo en la preferencia por los juguetes neutrales de género, como libros ilustrados o peluches. Presumiblemente, los monos no tienen un concepto previo de juguetes de “niño” o “niña”. Los monos rhesus jóvenes también muestran preferencias de juguete similares.
¿Qué subyace entonces a la diferencia de sexo en la preferencia por juguetes? Es posible que ciertos atributos de los juguetes (u objetos) atraigan ya sea a niños o niñas. Los juguetes que atraen a los niños o a los monos vervet o rhesus masculinos, en este caso, una pelota o un carro de juguete, son objetos que se pueden mover activamente por el espacio, juguetes que se pueden incorporar en el juego activo, áspero y de caída. El atractivo de los juguetes que prefieren las niñas o las hembras de monos vervet parece estar basado en el color. El rosa y el rojo (los colores de la muñeca y la olla) pueden provocar la atención a los bebés.
La sociedad puede reforzar esas respuestas estereotipadas a los juguetes típicos de género. Las diferencias de sexo en las preferencias de juguetes surgen a los 12 o 24 meses de edad y parecen fijas a los 36 meses de edad, pero ¿las diferencias de sexo en la preferencia de juguete están presentes durante el primer año de vida? Es difícil preguntar a los bebés preverbales qué prefieren, pero en estudios donde los investigadores examinaron la cantidad de tiempo que los bebés miraron diferentes juguetes, los datos de seguimiento ocular indican que bebés de tan solo tres meses mostraron diferencias de sexo en las preferencias de juguetes; las niñas preferían muñecas, mientras que los niños camiones preferidos. Otro resultado que sugiere, pero no prueba, que las hormonas están involucradas en las preferencias de los juguetes es la observación de que las niñas diagnosticadas con hiperplasia suprarrenal congénita (CAH), cuyas glándulas suprarrenales producen cantidades variables de andrógenos temprano en la vida, jugaban con juguetes masculinos con mayor frecuencia que las niñas sin CAH. Además, se observó una relación dosis-respuesta entre la extensión del trastorno (es decir, el grado de exposición a andrógenos fetales) y el grado de masculinización del comportamiento lúdico. ¿Son las diferencias de sexo en las preferencias de juguete o la actividad lúdica, por ejemplo, las consecuencias inevitables de los entornos hormonales diferenciales de niños y niñas, o estas diferencias son impuestas por prácticas y creencias culturales? ¿Estas diferencias son el resultado de recibir juguetes específicos de género desde una edad temprana, o son estas diferencias alguna combinación de factores hormonales y culturales? Nuevamente, estas son preguntas difíciles de desentrañar en las personas.
Comportamientos Agresivos
Obsérvese que en esta sección “comportamiento agresivo” se refiere específicamente a la expresión de agresión física (posiblemente el único tipo de agresión que se puede estudiar tanto en animales como en humanos). Cuando la agresión se define de manera más amplia para incluir la agresión relacional (como el acoso no físico, los chismes y la humillación), los hombres y las mujeres humanos muestran niveles generales de agresión mucho más similares (Swearer Napolitano, 2008).
La posibilidad de comportamiento agresivo existe siempre que los intereses de dos o más individuos están en conflicto (Nelson, 2006). Es más probable que surjan conflictos por recursos limitados como territorios, alimentos y compañeros. Una interacción social decide qué animal obtiene acceso al recurso impugnado. En muchos casos, una postura sumisa o gesto por parte de un animal evita la necesidad de un combate real sobre un recurso. Los animales también pueden participar en exhibiciones de amenazas o combates ritualizados en los que se determine el dominio pero no se inflija ningún daño físico.
Existe evidencia circunstancial abrumadora de que los andrógenos median el comportamiento agresivo en muchas especies. En primer lugar, las variaciones estacionales en las concentraciones plasmáticas de testosterona y las variaciones estacionales en la agresión coinciden Por ejemplo, la incidencia de comportamiento agresivo alcanza su punto máximo para los ciervos machos en otoño, cuando están secretando altos niveles de testosterona. Segundo, las conductas agresivas aumentan en el momento de la pubertad, cuando los testículos se vuelven activos y aumentan las concentraciones sanguíneas de andrógenos. Los venados juveniles no participan en los combates durante la época de apareamiento. Tercero, en cualquier especie dada, los machos son generalmente más agresivos que las hembras. Esto es ciertamente cierto en el caso de los ciervos; en relación con los ciervos, las hembras rara vez muestran un comportamiento agresivo, y sus raros actos agresivos son cualitativamente diferentes del comportamiento agresivo de los machos. Finalmente, la castración (extirpación de los testículos) generalmente reduce la agresión en los hombres, y la terapia de reemplazo de testosterona restaura la agresión a los niveles previos a la castración. Existen algunas excepciones interesantes a estas observaciones generales que están fuera del alcance de esta sección.
Como se mencionó, los machos son generalmente más agresivos que las hembras. Ciertamente, los machos humanos son mucho más agresivos físicamente que las hembras. Muchos más hombres que mujeres son condenados por delitos violentos en Norteamérica. Las diferencias de sexo en la agresividad humana aparecen muy tempranas. A todas las edades a lo largo de los años escolares, muchos más niños que niñas inician agresiones físicas. Casi todos reconocerán la existencia de esta diferencia de sexo, pero asignar una causa a las diferencias de sexo conductual en humanos siempre provoca mucho debate. Es posible que los niños sean más agresivos que las niñas porque los andrógenos promueven un comportamiento agresivo y los niños tienen mayores concentraciones de andrógenos en sangre que las niñas. Es posible que los niños y niñas difieran en su agresividad porque los cerebros de los niños están expuestos a los andrógenos prenatalmente y el “cableado” de sus cerebros se organiza así de una manera que facilita la expresión de la agresión (Figura\(\PageIndex{6}\)). También es posible que los niños sean alentados y las niñas sean desalentadas por la familia, compañeros u otras personas de actuar de manera agresiva. Estas tres hipótesis no son mutuamente excluyentes, pero es extremadamente difícil discriminar entre ellas para dar cuenta de las diferencias de sexo en la agresividad humana.

¿Qué tipo de estudios serían necesarios para evaluar estas hipótesis? Por lo general, es difícil separar las influencias del ambiente y la fisiología en el desarrollo del comportamiento en los humanos. Por ejemplo, niños y niñas difieren en su juego rudo a una edad muy temprana, lo que sugiere una influencia fisiológica temprana en la agresión. Este patrón de comportamiento de juego más físico se observa en otras especies, incluyendo primates no humanos, ratas y perros. ¿La diferencia en la frecuencia de juego rudo entre niños y niñas se debe a factores biológicos asociados con ser masculino o femenino, o se debe a expectativas culturales y aprendizaje? Los padres interactúan con su descendencia masculina y femenina de manera diferente; suelen jugar más bruscamente con bebés varones que con hembras, lo que sugiere que la diferencia de sexo en la agresividad se aprende parcialmente. Esta diferencia en el estilo de interacción parental es evidente para la primera semana de vida. Si hay una combinación de influencias biológicas y culturales que median la frecuencia del juego rudo y caída, entonces ¿qué proporción de la variación entre los sexos se debe a factores biológicos y qué proporción se debe a las influencias sociales? Es importante destacar, ¿es apropiado hablar de diferencias de sexo “normales” cuando estos rasgos prácticamente siempre se organizan a lo largo de un continuo en lugar de en categorías discretas? Debido a estas complejidades en los factores que influyen en el comportamiento humano, se ha perseguido el estudio de los efectos hormonales sobre el comportamiento diferenciado por sexo en animales no humanos, donde es más fácil mantener las influencias ambientales relativamente constantes.
Con el modelo animal apropiado, podemos abordar las preguntas planteadas anteriormente: ¿La diferencia de sexo en la agresión se debe a mayores concentraciones de andrógenos en la sangre adulta en los machos que en las hembras, o son los machos más agresivos que las hembras porque sus cerebros están organizados de manera diferente por hormonas perinatales? ¿Los machos suelen ser más agresivos que las hembras debido a una interacción de las concentraciones tempranas y actuales de andrógenos en sangre? Si los ratones machos son castrados antes de su sexto día de vida, luego tratados con testosterona en la edad adulta, muestran bajos niveles de agresión. De manera similar, los ratones hembra ovariectomizados antes de su sexto día pero a los que se les administraron andrógenos en la edad adulta no expresan niveles de agresión similares a los machos. El tratamiento de machos o hembras perinatalmente gonadectomizados con testosterona antes de su sexto día de vida y también en la edad adulta da como resultado un nivel de agresión similar al observado en ratones machos típicos. Así, en ratones, la proclividad para que los machos actúen de manera más agresiva que las hembras está organizada perinatalmente por los andrógenos pero también requiere la presencia de andrógenos después de la pubertad para que se expresen plenamente. En otras palabras, la agresión en ratones machos está organizada y activada por los andrógenos. La exposición a la testosterona en la edad adulta sin organización previa del cerebro por las hormonas esteroides no evoca niveles típicos de agresión masculina. El control hormonal del comportamiento agresivo en ratones es así similar a la mediación hormonal del comportamiento de apareamiento masculino heterosexual en otras especies de roedores. El comportamiento agresivo es igualmente organizado y activado por andrógenos en muchas especies, incluyendo ratas, hámsters, ratones de campo, perros y posiblemente algunas especies de primates.
Interpretaciones evolutivas de la conducta reproductiva 
La psicología evolutiva conecta los principios evolutivos con la psicología moderna y se centra principalmente en las adaptaciones psicológicas: cambios en la forma de pensar para mejorar nuestra supervivencia. (Consulte el Capítulo 3 para obtener más información sobre la evolución.) Esta sección describe una de las principales teorías psicológicas evolutivas que es relevante para los comportamientos reproductivos -la teoría de estrategias sexuales- que describe la psicología de las estrategias de apareamiento humano y las formas en que mujeres y hombres difieren en esas estrategias. Adicionalmente, se proporciona un ejemplo de otra teoría psicológica evolutiva -teoría de gestión de errores- que es relevante para el apareamiento humano.
Teoría de Estrategias Sexuales
La teoría de estrategias sexuales se basa en la teoría de la selección sexual, la evolución de las características debido a la ventaja de apareamiento más que a la ventaja de supervivencia, como los rasgos que hacen que un individuo sea más atractivo para el sexo opuesto, y la competencia entre miembros del mismo sexo. Propone que los humanos han desarrollado una lista de diferentes estrategias de apareamiento, tanto a corto como a largo plazo, que varían según la cultura, el contexto social, la influencia parental y el valor de la pareja personal (deseabilidad en el “mercado de apareamiento”). Inicialmente se enfocó en las diferencias entre hombres y mujeres en las preferencias y estrategias de apareamiento (Buss & Schmitt, 1993), comenzando por observar la mínima inversión parental necesaria para producir un hijo. Para las mujeres, incluso la inversión mínima es significativa: después de quedar embarazadas, tienen que llevar a ese niño durante nueve meses dentro de ellas (Figura\(\PageIndex{7}\)). Para los hombres, en cambio, la inversión mínima para producir el mismo hijo es considerablemente más pequeña, simplemente el acto sexual.

Estas diferencias en la inversión parental tienen un enorme impacto en las estrategias sexuales. Para una mujer, los riesgos asociados a tomar una mala elección de apareamiento son altos. Podría quedar embarazada de un hombre que no ayudará a mantenerla a ella y a sus hijos, o que podría tener genes de mala calidad. Y debido a que las apuestas son mayores para una mujer, las sabias decisiones de apareamiento para ella son mucho más valiosas. Si bien esta discusión no pretende de ninguna manera refrendar el “mal comportamiento”, para los hombres, biológicamente hablando, la necesidad de enfocarse en tomar decisiones sabias de apareamiento no es tan importante. Es decir, a diferencia de las mujeres, los hombres 1) biológicamente no tienen al niño creciendo dentro de ellos durante nueve meses, y 2) no tienen una expectativa cultural tan alta para criar al niño. Esta lógica lleva a un poderoso conjunto de predicciones: En el apareamiento a corto plazo, las mujeres probablemente serán más selectivas que los hombres (porque los costos de quedar embarazada son muy altos), mientras que los hombres, en promedio, probablemente participarán en actividades sexuales más casuales (porque este costo se ve muy disminuido). Debido a esto, los hombres a veces engañan a las mujeres sobre sus intenciones a largo plazo en beneficio del sexo a corto plazo, y los hombres tienen más probabilidades que las mujeres de bajar sus estándares de apareamiento para situaciones de apareamiento a corto plazo.
Un extenso cuerpo de evidencia empírica respalda estas predicciones y relacionadas (Buss & Schmitt, 2011). Los hombres expresan el deseo de tener un mayor número de parejas sexuales que las mujeres. Dejan que transcurra menos tiempo antes de buscar sexo. Están más dispuestos a dar su consentimiento para tener relaciones sexuales con extraños y es menos probable que requieran una participación emocional con sus parejas sexuales. Tienen fantasías sexuales más frecuentes y fantasean con una mayor variedad de parejas sexuales. Es más probable que se arrepientan de oportunidades sexuales perdidas. Y bajan sus estándares en el apareamiento a corto plazo, mostrando una disposición para aparearse con una mayor variedad de mujeres siempre y cuando los costos y riesgos sean bajos.
No obstante, también es importante señalar que en situaciones en las que tanto el hombre como la mujer están interesados en el apareamiento a largo plazo, ambos sexos tienden a invertir sustancialmente en la relación y en sus hijos. En estos casos, la teoría predice que ambos sexos serán extremadamente exigentes a la hora de perseguir una estrategia de apareamiento a largo plazo. Gran parte de la investigación empírica apoya esta predicción, también. De hecho, las cualidades que generalmente buscan mujeres y hombres al elegir parejas a largo plazo son muy similares: ambos quieren parejas que sean inteligentes, amables, comprensivas, saludables, confiables, honestas, leales, amorosas y adaptables.
Sin embargo, las mujeres y los hombres difieren en sus preferencias por algunas cualidades clave en el apareamiento a largo plazo, debido a problemas de adaptación algo distintos. Las mujeres modernas han heredado el rasgo evolutivo de desear parejas que posean recursos, tengan cualidades vinculadas con la adquisición de recursos (por ejemplo, ambición, riqueza, laboriosidad) y estén dispuestas a compartir esos recursos con ellas. Por otro lado, los hombres desean más fuertemente la juventud y la salud en las mujeres, ya que ambos son señales de fertilidad. Estas diferencias masculinas y femeninas son universales en los humanos. Se documentaron por primera vez en 37 culturas diferentes, desde Australia hasta Zambia (Buss, 1989), y han sido replicadas por decenas de investigadores en docenas de culturas adicionales (para resúmenes, ver Buss, 2012).
Como sabemos, sin embargo, solo porque tenemos estas preferencias de apareamiento (por ejemplo, hombres con recursos; mujeres fértiles), las personas no siempre obtienen lo que quieren. Hay un sinnúmero de otros factores que influyen a quienes las personas finalmente seleccionan como su pareja. Por ejemplo, la proporción de sexos (el porcentaje de hombres a mujeres en el grupo de apareamiento), las prácticas culturales (como los matrimonios concertados, que inhiben la libertad de los individuos para actuar sobre sus estrategias de apareamiento preferidas), las estrategias de los demás (por ejemplo, si todos los demás persiguen relaciones sexuales a corto plazo, es más difícil perseguir una estrategia de apareamiento a largo plazo), y muchos otros influyen en quienes seleccionamos como nuestros compañeros.
La teoría de las estrategias sexuales, anclada en la teoría de la selección sexual, predice similitudes y diferencias específicas en las preferencias y estrategias de apareamiento de hombres y mujeres. Ya sea que busquemos relaciones a corto o largo plazo, muchos factores de personalidad, sociales, culturales y ecológicos influirán en quiénes serán nuestros socios.
Teoría de Gestión de Errores
La forma en que pensamos, tomamos decisiones y evaluamos situaciones inciertas (teoría del manejo de errores) también se ha utilizado para predecir sesgos adaptativos en el dominio del apareamiento. Considera algo tan sencillo como una sonrisa. En un caso, una sonrisa de una pareja potencial podría ser una señal de interés sexual o romántico. Por otro lado, puede que solo señale amabilidad. Debido a los costos para los hombres de perderse oportunidades de reproducción, la teoría de la gestión de errores predice que los hombres tienen un sesgo de sobrepercepción sexual: a menudo malinterpretan el interés sexual de una mujer, cuando en realidad es solo una sonrisa amistosa o un toque. En el dominio del apareamiento, el sesgo de sobrepercepción sexual es uno de los fenómenos mejor documentados. Se ha demostrado en estudios en los que hombres y mujeres calificaron el interés sexual entre las personas en fotografías e interacciones grabadas en video. Además, se ha mostrado en el laboratorio con participantes participando en “citas rápidas” reales, donde los hombres interpretan el interés sexual de las mujeres con más frecuencia de lo que las mujeres realmente lo pretendían (Perilloux, Easton, & Buss, 2012). En definitiva, la teoría del manejo de errores predice que los hombres, más que las mujeres, sobreinferirán el interés sexual basándose en señales mínimas, y la investigación empírica confirma este sesgo de apareamiento adaptativo.
Resumen
Los humanos, como muchos animales, son sexualmente dimórficos en el tamaño y la forma de sus cuerpos, su fisiología, y para nuestros propósitos, su comportamiento. Las niñas generalmente sobresalen en habilidades verbales, mientras que los niños generalmente tienen mejores habilidades visuoespaciales. Los niños tienen más probabilidades de padecer dislexia, tartamudeo, autismo y esquizofrenia, mientras que la mayoría de los casos de anorexia nerviosa involucran a mujeres jóvenes. Las niñas se involucran en comportamientos de crianza con más frecuencia, y los niños generalmente participan en juegos más rudos y caídas. Una diferencia de sexo conductual que persiste hasta la edad adulta son los niveles más altos de agresión física exhibidos por niños y hombres.
En ratas, el núcleo sexualmente dimórfico del área preóptica en el hipotálamo (SDN-POA) es mucho más grande en machos que hembras. Una rata hembra tratada con testosterona cuando era recién nacida desarrolla un SDN-POA similar en tamaño a una rata macho típica. En humanos, se encontró que una zona cerebral similar, el tercer núcleo intersticial del hipotálamo anterior (INAH-3), era mayor en hombres heterosexuales que en hombres homosexuales (cuyo INAH-3 era similar en tamaño al de las mujeres). Sin embargo, la replicación fuera del laboratorio inicial no alcanzó significancia estadística. Los estudios de imágenes cerebrales han reportado diversas diferencias en regiones corticales y estructuras subcorticales, pero un escrutinio más detenido a veces hace que estas diferencias de sexo no sean importantes, y se debe tener precaución al inferir la causalidad. Un estudio de más de 1400 resonancias magnéticas de cerebros humanos encontró que al tomar en cuenta todo el cerebro, es imposible categorizar los cerebros en formas masculinas y femeninas. Hubo una amplia superposición de características “masculinas” y “femeninas” en los cerebros humanos y era muy raro tener consistencia interna en un cerebro específico. Estos hallazgos también apoyan la noción de que la masculinización y feminización de las áreas cerebrales son dos procesos separados que progresan de forma independiente, permitiendo una variación considerable entre los individuos.
También se han reportado diferencias de sexo en una serie de funciones sensoriales y cognitivas, pero estas diferencias en la capacidad son leves. Además, hay más variación dentro de cada sexo que entre los sexos para la mayoría de las habilidades cognitivas. Un análisis de los rasgos de personalidad, actitudes, intereses y comportamientos de más de 5.500 individuos reflejó los hallazgos de la RM (gran superposición entre hombres y mujeres y poca consistencia interna dentro del mismo individuo). Es razonable concluir que en general, los hombres y mujeres humanos son más similares de lo que son diferentes, tanto en lo que respecta a las características cerebrales como a los rasgos psicológicos.
Los niños generalmente prefieren juguetes como camiones y pelotas y las niñas generalmente prefieren juguetes como muñecas. Es posible que las hormonas den forma al desarrollo del cerebro de un niño para preferir ciertos tipos de juguetes o estilos de comportamiento de juego, pero los niños también aprenden qué tipos de juguetes y qué estilos de juego son apropiados a su género. Los estudios de monos vervet y monos rhesus encuentran que las hembras prefieren los juguetes típicos de las niñas, y los machos prefieren los juguetes típicos de los niños, sin diferencias de sexo en la preferencia por los juguetes neutros de género. Si bien la sociedad probablemente refuerza las respuestas estereotipadas a los juguetes típicos de género en niños humanos, las diferencias de sexo en las preferencias de juguetes surgen a los 12 o 24 meses de edad y parecen fijas a los 36 meses de edad. Los estudios que utilizan seguimiento ocular indican que los bebés de tan solo tres meses muestran diferencias estereotipadas de sexo en las preferencias de juguete. Adicionalmente, en una relación dosis-respuesta, las niñas diagnosticadas con hiperplasia suprarrenal congénita (CAH) jugaron con juguetes masculinos con más frecuencia que las niñas sin HCA, lo que sugiere que las hormonas prenatales pueden afectar las preferencias posteriores de los juguetes.
El comportamiento agresivo puede ocurrir cuando dos (o más) individuos están en conflicto, típicamente sobre recursos limitados como territorios, comida y compañeros. La evidencia sugiere fuertemente que los andrógenos median el comportamiento agresivo en muchas especies. Las variaciones estacionales en los niveles de testosterona y las variaciones en la agresión coinciden; los comportamientos agresivos aumentan en la pubertad (cuando los testículos se vuelven activos y aumentan los andrógenos); en cualquier especie dada, los machos son generalmente más agresivos que las hembras; y la castración (extirpación de los testículos) típicamente reduce la agresión en varones, mientras que la terapia de reemplazo de testosterona restaura la agresión. Los machos humanos son mucho más agresivos físicamente que las hembras, y estas diferencias de sexo aparecen muy tempranas. Es posible que los niños sean más agresivos que las niñas porque los andrógenos promueven un comportamiento agresivo, podría deberse a la exposición a andrógenos prenatalmente, o se puede alentar a los niños a actuar de manera agresiva. Estas tres hipótesis no son mutuamente excluyentes, pero es extremadamente difícil discriminar entre ellas para dar cuenta de las diferencias de sexo en la agresividad humana. En muchas especies animales (ratones, ratas, hámsters, ratones de campo, perros y posiblemente algunos primates), el comportamiento agresivo es organizado y activado por los andrógenos.
La teoría de estrategias sexuales describe la psicología de las estrategias de apareamiento humano y las formas en que mujeres y hombres difieren en esas estrategias. Debido a que las diferencias en la inversión de los padres son tan grandes entre hombres y mujeres- los hombres tienen poca inversión necesaria y en consecuencia no necesitan tomar decisiones sabias, mientras que las decisiones de apareamiento para las mujeres son mucho más importantes (ya que se requiere un mínimo de ninguno meses llevando al niño) - se pronostican las mujeres para ser más escogidos que los hombres en el apareamiento a corto plazo. Así, los hombres a veces engañan a las mujeres en beneficio del sexo a corto plazo, y los hombres tienen más probabilidades que las mujeres de bajar sus estándares de apareamiento para situaciones de apareamiento a corto plazo. Sin embargo, a la hora de elegir parejas a largo plazo, las cualidades que buscan mujeres y hombres son muy similares: parejas inteligentes, amables, comprensivas, saludables, confiables, honestas, leales, amorosas y adaptables. Sin embargo, hay algunas diferencias universales en la preferencia de pareja: las mujeres desean parejas que posean recursos (y estén dispuestas a compartirlos), y los hombres desean más fuertemente la juventud y la salud en las mujeres (ya que ambas son señales de fertilidad).
Referencias
Nota: Estas referencias son específicamente las agregadas al contenido de esta página por Naomi Bahm y no incluyen citas de las fuentes externas utilizadas. Consulte las atribuciones de texto para localizar citas de artículos de otras fuentes.
Bailey, J.M., Vasey, P.L., Diamond, L.M., Breedlove, S.M., Vilain, E., & Eppprecht, M. (2016). Orientación sexual, polémica y ciencia. La ciencia psicológica en el interés público, 17 (2), 45-101. doi: 10.1177/1529100616637616
Daly. M., & Wilson, M. (1996). Psicología evolutiva y conflicto conyugal. En Buss, D.M., & Malamuth, N.M. (editores), Sexo, Poder, Conflicto: Perspectivas Evolutivas y Feministas. Prensa de la Universidad de Oxford.
Goldstein, J., Seidman, L.J., Horton, N.J., Makris, N., Kennedy, D.N., Caviness, V.S. Jr., Faraone, S.V., & Tsuang M.T. (2001). Dimorfismo sexual normal del cerebro humano adulto evaluado por resonancia magnética in vivo. Corteza Cerebral, 11, 490-497.
Jaffe, K.E. (2019). Capítulo 6: Comportamiento de Primates y Ecología. En Shook, B., Nelson, K., Aguilera, K.& Braff, L. (editores), Exploraciones: una invitación abierta a la antropología biológica. Asociación Antropológica Americana, licenciada CC BY-NC 4.0 Internacional, ISBN 978-1-931303-63-7
Joel, D., Berman, Z., Tavor, I., Wexler, N., Gaber, O., Stein, Y., Shefi, N., Pool, J., Urchs, S., Margulies, D.S., Liem, F., Hänggi, J., Jäncke, L. & Assaf, Y. (2015). Sexo más allá de los genitales: El mosaico del cerebro humano. PNAS, 112 (50), 15468-15473. https://doi.org/10.1073/pnas.1509654112
Larsen, C.S. (2003). ¿Igualdad para los sexos en la evolución humana? Dimorfismo sexual homínido temprano e implicaciones para los sistemas de apareamiento y el comportamiento social. Actas de la Academia Nacional de Ciencias de los Estados Unidos de América, 100 (16), 9103—9104. https://www.pnas.org/doi/10.1073/pnas.1633678100
LeVay, S. (1991). Una diferencia en la estructura hipotalámica entre hombres heterosexuales y homosexuales. Ciencia, 253 (5023), 1034-1037. doi: 10.1126/ciencia.1887219
Luders, E., Toga, A.W., & Thompson, P.M. (2014) Por qué importa el tamaño: Las diferencias en el volumen cerebral explican las diferencias aparentes de sexo en la anatomía callosal. El dimorfismo sexual del cuerpo calloso. NeuroImage, 84, 820-824. http://dx.doi.org/10.1016/j.neuroimage.2013.09.040
McDowell, M.A., Fryar, C.D., Ogden, C.L., & Flegal, K.M. (2008). Datos antropométricos de referencia para niños y adultos: Estados Unidos, 2003-2006. Informes Nacionales de Estadísticas de Salud, 10 (22 de octubre de 2008), 1-48. https://www.cdc.gov/nchs/data/nhsr/nhsr010.pdf
Swearer Napolitano, S.M. (2008) Agresión relacional: No sólo un tema femenino. Revista de Psicología Escolar, 46, 611-616. doi: 10.1016/j.jsp.2008.08.001 [disponible en https://digitalcommons.unl.edu/edpsychpapers/142/]
van Eijk, L., Zhu, D., Couvy-Duchesne B., Strike, L.T., Lee, A.J., Hansell, N.K., Thompson, P.M., de Zubicaray, G.I., McMahon, K.L., Wright, M.J., & Zietsch, B.P. (2021) ¿Las diferencias de sexo en la estructura del cerebro humano están asociadas con diferencias sexuales en el comportamiento? Ciencia Psicológica, 32 (8), 1183-1197. doi: 10.1177/0956797621996664
Recursos adicionales
Web: Principal organismo científico internacional para el estudio de la evolución y el comportamiento humano, HBES
Web: Libros y entrevistas con David Buss
https://labs.la.utexas.edu/buss/books/
Web: Publicaciones de Buss y colegas
https://labs.la.utexas.edu/buss/publications/
Artículo de revista: Fundadores de la Psicología Evolutiva
Atribuciones
- Cifras:
- Gallo y gallina de John Cudworth, licenciado CC BY-NC 2.0, encontrado en NOBA Hormonas y Comportamiento por Randy Nelsen
- Foto izquierda: Gibbons de mano blanca de Cliff de Arlington, Virginia, Estados Unidos, CC BY 2.0 vía Wikimedia Commons; Imagen del medio: Humanos extraídos de la placa Pioneer por la NASA; vectores de Mysid, Dominio público, vía Wikimedia Commons; Foto derecha: Gorilas de montaña por Joachim Huber, CC BY-SA 2.0, vía Wikimedia Commons
- SDN-POA, sin atribución o información de licencia, encontrada en NOBA Hormonas y Comportamiento por Randy Nelsen
- Gráfico de superposición de diferencias de sexo, sin atribución ni información de licencia, encontrada en NOBA Hormonas y Comportamiento por Randy Nelsen
- Izquierda: Hermano y hermana, por Amanda Westmont, https://goo.gl/ntS5qx, licenciada CC BY-NC-SA 2.0, encontrada en NOBA Desarrollo Social y de la Personalidad en la Infancia por Ross Thompson; Derecha: Niña sentada, licenciada CC0 Public Domain, encontrada en NOBA Hormonas y Comportamiento por Randy Nelsen
- Expresión agresiva por Riccardo Cuppini, CC BY-NC-ND 2.0,[1] encontrada en NOBA Hormonas y Comportamiento por Randy Nelsen
- Mujer embarazada, con licencia CC0 Public Domain, encontrada en NOBA Teorías Evolutivas en Psicología por David Buss
- Texto adaptado de:
- Hormonas y comportamiento por Randy J. Nelson, con licencia CC BY-NC-SA 4.0 vía Noba Project.
- Teorías Evolutivas en Psicología por David M. Buss, licenciado CC BY-NC-SA 4.0 vía Noba Project.
- Cambios: Texto (e imágenes) de dos fuentes anteriores reconstruido con algunas modificaciones, transiciones y contenido adicional (particularmente en las secciones Nota Preliminar, Introducción a las Diferencias de Sexo, Diferencias de Sexo en el Cerebro y Diferencias de Sexo en Sistemas Sensoriales y Cognición) agregadas por Naomi I. Gribneau Bahm, PhD., Profesor de Psicología en Cosumnes River College, Sacramento, CA.