
14.5: Mecanismos cerebrales e inteligencia
- Page ID
- 148242
- Describir las estructuras cerebrales incluidas en el modelo P-FIT de la biología de la inteligencia.
- Describir la Red de Modo Predeterminado (DMN) y su relación con la biología de la inteligencia.
- Discutir el tipo de atención que probablemente implica cuando alguien está tomando una prueba de I.Q. y cómo esto se relaciona con la interpretación de estudios de imágenes cerebrales de las redes cerebrales involucradas en la inteligencia humana.
Visión general
Según Hearne, et al. (2016, p.1), “La inteligencia humana puede definirse ampliamente como la capacidad de comprender ideas complejas, adaptarse eficazmente al entorno y participar en razonamientos complejos 1. Las medidas de inteligencia pueden estar relacionadas con el desempeño en prácticamente cualquier tarea cognitiva, desde la discriminación sensorial 2 hasta tareas cognitivas desafiantes como la identificación de patrones en la prueba de Matrices Progresivas del Cuervo 3. Es importante destacar que los puntajes en las pruebas de inteligencia pueden predecir con precisión diversos resultados de vida, incluyendo el éxito académico 4, el desempeño laboral 5 y la morbilidad y mortalidad de adultos 6 A pesar de la importancia adaptativa de la inteligencia en la vida humana, no se comprenden bien los mecanismos cerebrales involucrados en la inteligencia. Sin embargo, la investigación deja cada vez más claro que funciones complejas como la cognición y la inteligencia involucran muchas áreas cerebrales que actúan juntas en redes neuronales interconectadas compuestas por múltiples áreas cerebrales. Además, las interacciones entre diferentes redes de áreas cerebrales también son importantes si queremos entender cómo el cerebro crea cognición e inteligencia. La resonancia magnética funcional (fMRI) se ha utilizado para investigar la relación entre las diferencias individuales en la inteligencia y la actividad cerebral durante actividades cognitivas como la memoria de trabajo y el razonamiento. Como se discutió en módulos anteriores de este capítulo, los estudios de imágenes cerebrales implican interacciones neuronales en una red fronto-parietal subyacente a muchas de las funciones asociadas con la inteligencia humana. Estas observaciones han dado como resultado la influyente Teoría de la Integración Parieto-Frontal de la Inteligencia (P-FIT) que identifica una serie de estructuras cerebrales interactuantes asociadas con diferencias individuales en la inteligencia humana, incluidos los lóbulos frontal y parietal y otras estructuras. Investigaciones adicionales (Hearne, et al., 2016) muestran que las diferencias individuales en inteligencia entre las personas también están asociadas con el grado de interacción neuronal entre la red fronto-parietal y la red de modo predeterminado. La mayor conectividad entre estas dos redes neuronales cuando el cerebro está en reposo (es decir, en ausencia de cualquier tarea cognitiva específica) se correlaciona con puntuaciones de inteligencia más altas en individuos en comparación con aquellos con menor conectividad entre las redes fronto-parietal y de modo predeterminado. También se deben considerar las interacciones con una red de atención dorsal (DAN) para una comprensión más completa de las complejas interacciones de red involucradas en la cognición y la inteligencia.
Mecanismos cerebrales e inteligencia
Debido a la complejidad y cantidad de factores que interactúan en la inteligencia humana, comprender los mecanismos cerebrales en la inteligencia humana es una tarea monumental para los investigadores. La siguiente figura que muestra el influyente modelo de inteligencia humana de Carroll es un recordatorio de la gran cantidad de habilidades de interacción involucradas. Los diversos factores y habilidades mostrados se derivan matemáticamente utilizando análisis factorial basado en el análisis de patrones de correlaciones en el desempeño de un gran número de personas en medidas de inteligencia como las pruebas estándar de I.Q. (e.g. Wechsler Adult Intelligence Scale, WAIS).
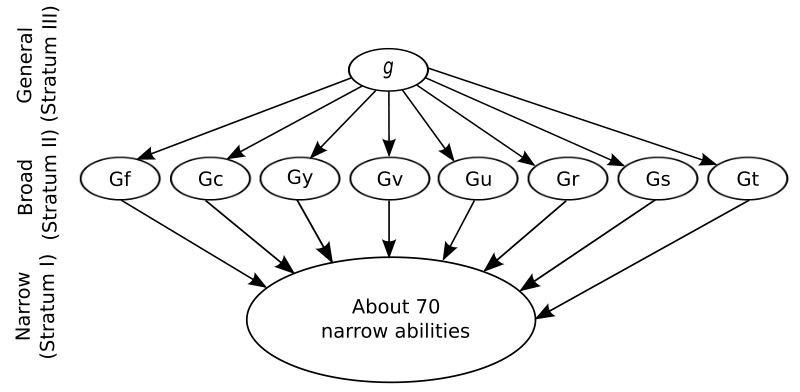
Figura\(\PageIndex{1}\): En la parte superior del modelo de inteligencia humana de tres estratos de Carroll se encuentra el factor g, la inteligencia general. Observe el gran número de habilidades del Estrato I que comprenden los factores en g y los factores g más específicos en el Estrato II. Abreviaturas: inteligencia fluida (Gf), inteligencia cristalizada (Gc), memoria general y aprendizaje (Gy), percepción visual amplia (Gv), percepción auditiva amplia (Gu), amplia capacidad de recuperación (Gr), amplia rapidez cognitiva (Gs) y velocidad de procesamiento (Gt). (Imagen y abreviaturas de Wikimedia Common; Archivo:Carroll tres stratum.svg; https://commons.wikimedia.org/wiki/F...ee_stratum.svg; por Victor Chmara; puesto a disposición bajo el dominio público universal Creative Commons CC0 1.0 Dedicación).
A pesar de esta complejidad, los neurocientíficos que utilizan métodos de imagen cerebral han identificado regiones cerebrales asociadas con diferencias en el coeficiente intelectual medido u otras medidas de inteligencia general. Como se mencionó anteriormente en este capítulo, Jung y Haier (2007) revisaron estudios de imágenes cerebrales e identificaron un conjunto de áreas cerebrales correlacionadas con diferencias individuales en inteligencia y razonamiento entre las personas. Llamaron a este conjunto de regiones cerebrales interconectadas la red fronto-parietal y denominaron a su modelo la Teoría de la Integración Parietal-Frontal (P-FIT). “El modelo P-FIT incluye, por áreas de Brodmann (BA): la corteza prefrontal dorsolateral (BAs 6, 9, 10, 45, 46, 47), el lóbulo parietal inferior (BA 39, 40) y superior (BA 7), el cingulado anterior (BA 32) y las regiones dentro los lóbulos temporal (BAs 21, 37) y occipital (BAs 18, 19). También están implicadas regiones de materia blanca (es decir, fasciculus arqueado)” (Jung y Haier, 2007, p. 135).
El fascículo arqueado interconecta regiones de la corteza temporal y parietal con la corteza frontal (ver Figura 14.8.2). Este haz de axones conecta las áreas de Wernicke y Broca, en lóbulos temporales y frontales, respectivamente, que están involucradas en la comprensión del lenguaje y la producción del lenguaje (véanse los módulos sobre el lenguaje más adelante en este capítulo). Sorprendentemente, esto no significa que la habilidad del lenguaje sea necesaria para la cognición y la inteligencia.
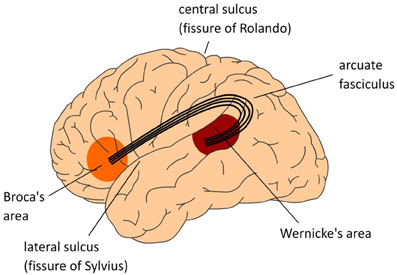
Figura\(\PageIndex{2}\): (Izquierda): El Fasciculo Arcuado. Vista lateral del hemisferio izquierdo. (Centro): La figura central también muestra una vista lateral del hemisferio izquierdo. Los números indican áreas de Brodmann (BA). Estas son áreas con diferencias en la citoarquitectónica (es decir, composición de tipos celulares). Las áreas de memoria se encuentran en la corteza temporal (en amarillo) incluyendo la circunvolución angular en la corteza parietal. Área de Broca (áreas Brodmann 44 y 45) y corteza adyacente (áreas Brodmann 47 y 6) en el lóbulo frontal involucrado en el lenguaje. Las operaciones de control reclutan otra parte del lóbulo frontal (en rosa), y la Corteza Cingulada Anterior (ACC; no mostrada en la figura central), así como áreas involucradas en la atención. (Derecha): Vista medial de la corteza cerebral que muestra giro mayor. (Imágenes de Wikimedia Commons; Imagen izquierda: Archivo:El modelo clásico de Wernicke-Lichtheim-Geschwind de la neurobiología del lenguaje fpsyg-04-00416-g001.jpg; https://commons.wikimedia.org/wiki/F...00416-g001.jpg; por Peter Hagoort; licenciado bajo la Creative Commons Atribución 3.0 Licencia Unported. Centro de imagen y subtítulo: Archivo:El modelo MUC (Memoria, Unificación, Control) del lenguaje fpsyg-04-00416-g002.jpg; https://commons.wikimedia.org/wiki/F...00416-g002.jpg; por Peter Hagoort; Hagoort P (2013) MUC (Memoria, Unificación, Control) y más allá. Frente. Psychol. 4:416. doi: 10.3389/fpsyg.2013.00416 http://journal.frontiersin.org/article/10.3389/fpsyg.2013.00416/full); licenciado bajo la licencia Creative Commons Attribution 3.0 Unported. Imagen derecha: Archivo:Superficie medial de la corteza cerebral - gyri.png; https://commons.wikimedia.org/wiki/F...tex_-_gyri.png; por Patric Hagmann et.al., Hagmann P, Cammoun L, Gigandet X, Meuli R, Honey CJ, et al. (2008) Mapping the Structural Core of Human Cerebral Cortex. PLoS Biol 6 (7): e159. doi:10.1371/journal.pbio.0060159 [1]; licenciado bajo la licencia Creative Commons Attribution 2.5 Genérica).
Por ejemplo, “los pacientes con daños incluso graves en la red del lenguaje pueden retener alta inteligencia y la capacidad de realizar tareas cognitivas desafiantes, como la aritmética (Varley et al., 2005) y el razonamiento causal (por ejemplo, Varley y Siegal, 2000; ver Fedorenko y Varley, 2016, para una revisión)” (Assem, et al., 2020 , p. 139). Este hallazgo sugiere fuertemente que las tareas cognitivas como la aritmética y el razonamiento causal no dependen del lenguaje o de las redes lingüísticas. Este hallazgo es consistente con la teoría de que el razonamiento causal es un componente de la inteligencia general evolucionada por la “internalización genética” de la causalidad, similitud y relaciones predictivas en la organización funcional del cerebro, y que la inteligencia general se encuentra en muchas especies animales no humanas a pesar de que les falta lenguaje (Koenigshofer, 2017).
Utilizando fMRI, Duncan (2010, 2013) ha identificado un patrón frontal y parietal de actividad neuronal asociado con diversas tareas cognitivas e inteligencia fluida general. Duncan llama a este patrón “multi-demanda” (MD) porque este patrón de activación es una “parte sobresaliente de la respuesta del cerebro a muchos tipos diferentes de desafío cognitivo. [T] su patrón de demanda múltiple (MD) se extiende sobre un conjunto específico de regiones en la corteza prefrontal y parietal, en particular: corteza en y alrededor de la parte posterior del surco frontal inferior (IFS), [córtex' en la insula anterior y la adyacente opérculo frontal (AI/FO), en el área motora pre-suplementaria y cingulado anterior dorsal adyacente (Presma/ACC), y [corteza] en y alrededor del surco intraparietal (IPS). A veces se observa una región más pequeña de actividad acompañante en la corteza prefrontal rostrolateral (RPFC)” (Duncan, 2010, p. 172). Se observa un patrón similar de activación en tareas asociadas a la inteligencia fluida general. Además, “datos recientes de lesiones sugieren que los déficits en la inteligencia de fluidos se asocian específicamente con el daño a la red MD (Duncan, 2010, p. 172).
Mediante el uso de fMRI, Gray, et al. (2003) encontraron que las diferencias individuales en la inteligencia fluida general (gF) se asociaron con diferencias en la activación de los mecanismos atencionales en la corteza prefrontal lateral y parietal durante una tarea de memoria de trabajo. Estos resultados son consistentes con el modelo P-FIT de inteligencia y sugieren que al menos algunas de las diferencias en la inteligencia medida entre las personas pueden deberse a diferencias en las redes cerebrales que median la atención.
Según Assem, et al. (2020, p. 131), “Una red de Demanda Múltiple Frontoparietal distribuida (MD) [Duncan, 2010, 2013] ha estado implicada durante mucho tiempo en el comportamiento inteligente, y su daño se ha asociado con menor inteligencia y dificultades en la resolución de problemas. [Se] se ha vinculado a nuestra capacidad de involucrarnos en comportamientos dirigidos por objetivos, resolver problemas novedosos y adquirir nuevas habilidades. El daño a esta red como resultado de un accidente cerebrovascular, degeneración o lesión en la cabeza conduce a habilidades ejecutivas más pobres (atención, memoria de trabajo y control inhibitorio) y menor inteligencia fluida (Glascher et al., 2010; Roca et al., 2010; Woolgar et al., 2010). y el funcionamiento aberrante de esta red ha sido el funcionamiento aberrante de esta red ha sido reportados en una variedad de trastornos neurológicos y psiquiátricos”.
Al enfatizar las interacciones entre diversas redes cerebrales en la inteligencia humana, Hearne, et al. (2016, p. 7) afirman: “Cómo el cerebro se autoreorganiza para lograr configuraciones óptimas de redes funcionales a través de individuos con diferentes niveles de inteligencia es una pregunta abierta. Trabajos recientes de neuroimagen han sugerido que la cooperación transitoria entre diferentes sistemas neuronales, incluyendo las redes fronto-parietal, cingulo-opercular y de modo predeterminado, es parte integral de tareas cognitivas complejas como el razonamiento 42 , 43, recolección de memoria 44 y rendimiento de memoria de trabajo 39, 45. Estudios futuros deben probar la noción de que las diferencias individuales en inteligencia se basan en reconfiguraciones dinámicas, específicas del contexto, de la actividad local y la conectividad dentro de un sistema difuso que comprende regiones fronto-parietal, cingulo-opercular y de modo predeterminado 46.”
Hearne, et al. (2016, p. 1) “revelaron una contribución novedosa de las interacciones a través de la red entre el modo predeterminado y las redes fronto-parietales a las diferencias individuales en la inteligencia en reposo (es decir, en ausencia de cualquier tarea cognitiva específica). Específicamente, [ellos] encontraron que una mayor conectividad en el estado de reposo se asoció con mayores puntajes de inteligencia”. La red de modo predeterminado incluye la corteza prefrontal medial, la corteza cingulada posterior y el lóbulo parietal inferior. Otras estructuras a veces consideradas parte de esta red son el lóbulo temporal medio y el precuneo.
¿Cuál es el significado de la conectividad entre redes en reposo, cuando no se realiza ninguna tarea cognitiva específica? La respuesta a esta pregunta refleja otro método utilizado por los neurocientíficos para comprender las relaciones funcionales y neuroanatómicas entre redes de áreas cerebrales interconectadas. Dixon et al. (2017) describen el método de esta manera: “La conectividad funcional del estado de reposo ha surgido como una herramienta poderosa y no invasiva para delinear la arquitectura de red funcional del cerebro humano”. En otras palabras, la conectividad entre redes cerebrales en reposo ayuda a los neurocientíficos a determinar cómo varias redes cerebrales están “conectadas” entre sí y cómo interactúan funcionalmente entre sí para producir cognición e inteligencia.
En esta luz, recuerde que Hearne, et al. (2016, p.1) encontraron que “una mayor conectividad en el estado de reposo [entre las redes de modo predeterminado y fronto-parietales] se asoció con mayores puntuaciones de inteligencia”. Para ayudarnos a entender por qué esto podría ser así, tenemos que saber algo sobre el papel sospechoso que juega la red en modo predeterminado en la cognición. Dixon, et al. (2017, p.632) lo explican de esta manera: “La red por defecto (DN) está involucrada en una variedad de procesos dirigidos internamente, incluyendo la autorreflexión, la memoria autobiográfica, la simulación de eventos futuros, el procesamiento conceptual y la cognición espontánea. y exhibe una activación disminuida. durante muchas tareas cognitivas que exigen atención perceptual externa”. El hallazgo de disminución de la activación durante tareas cognitivas que requieren atención a los aportes del entorno externo sugirió a algunos investigadores que podrían haber dos redes competitivas mutuamente inhibidoras, una para la reflexión interna, y la otra, una” tarea positiva” o “relacionada con tareas” para la atención al mundo externo. Sin embargo, investigaciones posteriores encontraron “coactivación y conectividad funcional positiva entre el DN y la red de control frontoparietal (FPCN) —un componente de la red “tarea positiva” ―durante algunas condiciones de tarea, incluyendo el deambular mental, el pensamiento espontáneo, la planeación autobiográfica del futuro, la creatividad, la memoria recuerdo, memoria de trabajo guiada por información no relacionada con la entrada perceptual actual, la memoria de trabajo social y la toma de decisiones semánticas. Por otro lado, los estudios generalmente han encontrado anticorrelación [el aumento de la actividad en una red se asocia con una disminución en la otra] entre el DN y otros componentes de la red “tarea positiva”, particularmente la red de atención dorsal (DAN)” (Dixon, et al., 2017, p. 633).
La “red de atención dorsal” (DAN) o “red frontoparietal” dirige la atención visual y los procesos de memoria a corto plazo. Además, esta red es distinta de una red de control cognitivo cingulo-opercular (CCN). Sin embargo, no se ha llegado a un consenso respecto a los componentes precisos del DAN. Sobre la base de estudios de fMRI basados en tareas y en estado de reposo, el DAN en humanos se define típicamente para incluir todas o algunas de las siguientes cuatro regiones: (1) surco intraparietal (IPS)/lóbulo parietal superior; (2) surco precentral superior (SPC) que contiene el homólogo de campos oculares frontales de primates; (3) surco precentral inferior (CIP), alternativamente conocido como unión frontal inferior; y (4) el complejo MT de área sensible al movimiento (MT). [Además,] las estructuras subcorticales, como el cólico superior y el pulvinar, a menudo están implicadas en funciones atencionales” y partes del cerebelo también están involucradas en el DAN” (Brissenden, et al., 2016, p. 6083-4).
Figura\(\PageIndex{3}\): (Izquierda): La Red de Modo Predeterminado, vistas de sección transversal media sagital y horizontal. (Derecha): Red de modo predeterminado contrastada con Red relacionada con tareas. Sobre un fondo verde, la red de modo predeterminado se resalta en colores cálidos (rojo y amarillo) y la red relacionada con tareas se resalta en colores fríos (azul y azul claro). Vistas superiores y mediales. Vistas inferiores, laterales. (Imágenes de Wikimedia Commons; Izquierda: Archivo:Modo predeterminado network-WRNMMC.jpg; https://commons.wikimedia.org/wiki/F...ork-WRNMMC.jpg; por John Graner, Departamento de Neuroimagen, Centro Nacional de Excelencia Intrepid, Centro Médico Militar Nacional Walter Reed; en el dominio público en Estados Unidos por tratarse de un trabajo preparado por un funcionario o empleado del Gobierno de los Estados Unidos como parte de los deberes oficiales de esa persona. Derecha: Archivo:Modo predeterminado y mapas relacionados con tareas para subjects.jpg saludable; https://commons.wikimedia.org/wiki/F...y_subjects.jpg; por Shim G, Oh JS, Jung WH, et al., Shim G, Oh JS, Jung WH, et al. Conectividad alterada en estado de reposo en sujetos con riesgo ultra alto de psicosis: un estudio de fMRI. Funciones conductuales y cerebrales: BBF. 2010; 6:58. doi:10.1186/1744-9081-6-58. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2959003/; Shim G, Oh JS, Jung WH, et al. Conectividad alterada en estado de reposo en sujetos con riesgo ultra alto de psicosis: un estudio de fMRI. Funciones conductuales y cerebrales: BBF. 2010; 6:58. doi:10.1186/1744-9081-6-58. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2959003/).
“[A] ttention es un mecanismo de selección que sirve para elegir una fuente particular de estimulación, un tren interno de pensamientos, o un curso de acción específico para el procesamiento prioritario. Sin embargo, en ciertas situaciones es necesaria la atención para supervisar la acción dirigida a objetivos. [y] es necesario para detectar errores y controlar el comportamiento en condiciones peligrosas y novedosas o no practicadas. Así, los mecanismos de atención también son centrales para la generación de conductas voluntarias, que a menudo implican la inhibición de las respuestas automáticas. [Además, la atención puede ser] principalmente impulsada por la estimulación externa o. [por] procesos endógenos como intenciones voluntarias o expectativas... [o] la atención también puede dirigirse a un objeto por su relevancia para nuestros objetivos actuales” (Rueda, et al., 2015, p.183). Adicionalmente, se debe mantener una atención sostenida en una tarea (como tomar una prueba de I.Q.) durante la distracción. La atención sostenida y enfocada está claramente involucrada en el desempeño de una persona en muchas medidas de inteligencia. Rueda, et al. (2015) identifican tres tipos de redes de atención: 1) alertar, como la atención a un ruido fuerte repentino (mediado por la norepinefrina); 2) orientación, desplazamiento de atención de una ubicación a otra, involucrando cortezas parietales y frontales y que usa acetilcolina; y 3) atención ejecutiva, atención requerida para lidiar con la distracción e información incongruente (conflictiva) especialmente si es incompatible con la tarea, meta o problema en cuestión; la atención ejecutiva involucra cortezas cíngulas anteriores y prefrontales, y se modula por los niveles de dopamina cerebral y serotonina.
La estrecha asociación de la atención con la capacidad de concentración durante la memoria de trabajo y la resolución de problemas plantea la posibilidad de una confusión entre la atención y la inteligencia en algunos de los estudios de imagen cerebral que apoyan el P-FIT y modelos similares de la biología de la inteligencia discutidos anteriormente. Es decir, las redes identificadas con inteligencia pueden llegar a definirse con mayor precisión como redes de atención. Si es así, parte de la investigación de imágenes cerebrales en inteligencia puede estar accediendo a redes atencionales, en lugar de redes de inteligencia. En este caso, las variaciones en el I.Q. asociadas al funcionamiento de estas redes pueden reflejar principalmente diferencias entre las personas en los mecanismos atencionales que luego afectan el desempeño en las pruebas I.Q. La atención y la memoria de trabajo, aunque ciertamente esenciales para la resolución de problemas, el razonamiento y la acción inteligente, no captan los propios procesos de razonamiento. Al interpretar la investigación en esta área, es importante conocer las características específicas de las tareas cognitivas en estudio. Por ejemplo, ¿algún estudio de imágenes en particular utiliza procedimientos de control que puedan separar los efectos de la atención y la memoria de trabajo de los propios procesos de razonamiento de la inteligencia? Puede resultar que esta separación sea tan difícil de lograr que, en la práctica real, el solapamiento entre las redes de atención y las redes de inteligencia puede ser difícil de desentrañar. Sin embargo, se han identificado redes atencionales como la red de atención dorsal (DAN) y una red de control cognitivo cingulo-opercular (CCN) (y otras mencionadas anteriormente), y la medida en que estas no se superponen a las redes asociadas a la inteligencia fluida general, los investigadores pueden ser más cerca de identificar las redes involucradas en el razonamiento y la inteligencia como distintas de los mecanismos de atención.
Mientras tanto, sí sabemos que las variaciones entre las personas en I.Q. y otras medidas de inteligencia se correlacionan con las diferencias en su actividad cerebral en la red fronto-parietal (como lo proponen el modelo P-FIT y la red MD de Duncan), y con los grados de conectividad funcional en reposo entre esta red y la red de modo por defecto. Sin embargo, se necesitan investigaciones adicionales para aclarar las razones subyacentes de estas correlaciones entre las medidas de inteligencia y los niveles de actividad en estas redes cerebrales y su conectividad.
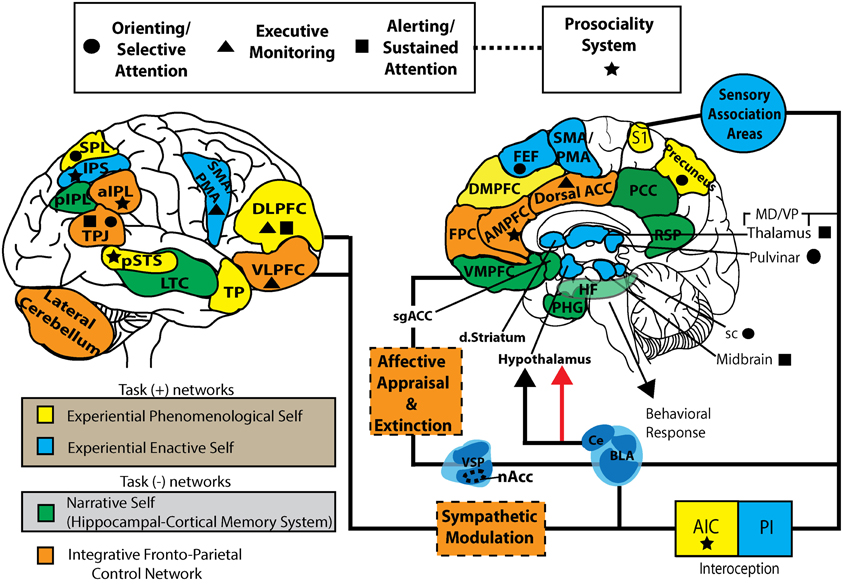
Figura\(\PageIndex{4}\): Un modelo de las complejas interacciones entre diversas áreas cerebrales involucradas en el control cognitivo de la conducta generando diversos modelos neuronales del yo inteligente en interacción con los mundos interno (mental) y externo (sensorial) (el subtítulo anterior del autor de este módulo, Kenneth A. Koenigshofer, Ph.D.). (El siguiente subtítulo es de los autores de esta imagen). Este modelo de trabajo representa una parcelación de tareas positivas (autoespecificantes: EPS y EES), negativas para tareas (NS) y redes integradoras de control fronto-parietal. EPS: yo fenomenológico experiencial; EES: yo vivencial enactivo; NS: yo narrativo; FPCN: red de control fronto-parietal; FEF: campos oculares frontales; DMPFC, corteza prefrontal dorsal-medial; AMPFC, corteza prefrontal medial anterior; VMPFC, corteza prefrontal ventromedial; PHG, giro parahipocampo; IC, formación hipocampal; RSP: corteza retrosplenial; PCC: corteza cingulada posterior; ACC dorsal, corteza cingulada anterior dorsal; DLPFC, corteza prefrontal dorsolateral; VLPFC, corteza prefrontal ventrolateral; TP, polo temporal, LTC, corteza temporal lateral; TPJ: unión temporoparietal; SPL, parietal superior lóbulo; pIPl, lóbulo parietal inferior posterior; AiPL, lóbulo parietal inferior anterior; NACC, núcleo accumbens; VSP, palidio ventrostriatal; dstriatum, estriado dorsal; S1, corteza somatosensorial primaria; AIC, corteza insular anterior; PIC: corteza insular posterior; SGACC, corteza cingulada anterior subgenual; VMPo, Núcleo posterior ventromedial; sc, colículo superior; BLA, amígdala basolateral; CE, núcleo central. (Imagen de Wikimedia Commons; Archivo:S-Art Mindfulness y brain1.jpg; https://commons.wikimedia.org/wiki/F...and_brain1.jpg; por Vago DR y Silbersweig DA; licenciado bajo la licencia Creative Commons Attribution 3.0 Unported).
Bases celulares de las diferencias en la inteligencia humana
La mayor parte de la investigación sobre la base neurológica de la inteligencia en humanos se ha centrado ya sea en 1) loci génicos asociados con diferencias individuales en la inteligencia o 2) se basa en el uso de imágenes cerebrales completas para identificar regiones cerebrales correlacionadas con diferencias individuales en inteligencia. Algunos estudios muestran genes comunes para las diferencias en el volumen cerebral y diferencias en la inteligencia, mientras que otros sugieren que los genes que facilitan el crecimiento de las neuronas están asociados con un coeficiente intelectual más alto (ver sección a continuación sobre genes e inteligencia). Los estudios de resonancia magnética funcional y estructural destacan correlaciones entre áreas específicas de la corteza, incluyendo lóbulos frontal y temporal, y medidas de g, inteligencia general (ver discusión de g en módulos anteriores en este capítulo). La corteza de asociación de orden superior en los lóbulos frontal y temporal contiene un gran número de neuronas piramidales con ramificaciones dendríticas grandes y complejas que podrían explicar diferencias individuales en el grosor cortical, la integración sináptica y quizás el coeficiente intelectual (Goriounova, et al., 2018). Estas neuronas y sus conexiones son “los principales bloques de construcción para la codificación, el procesamiento y el almacenamiento de información en el cerebro y dan lugar a la cognición (Salinas y Sejnowski, 2001). Dado su gran número en el neocórtex humano, incluso el más mínimo cambio en la eficiencia de la transferencia de información por las neuronas puede traducirse en grandes diferencias en la capacidad mental” (Goriounova, et al., 2018, p. e41715).
Como se muestra en la Figura 14.5.5 (abajo), Goriounova, et al. (2018) encontraron que las puntuaciones de CI se correlacionan positivamente con el grosor cortical del lóbulo temporal, la longitud de las dendritas neuronales piramidales y la complejidad de la ramificación dendrítica. “Así, las neuronas piramidales más grandes y complejas en el área de asociación temporal pueden contribuir en parte a una corteza más gruesa y vincularse a una inteligencia superior” (p. e41721).
Estos mismos investigadores también encontraron evidencia de que las neuronas piramidales del lóbulo temporal más grandes generaban potenciales de acción más rápido que las más pequeñas y que el inicio del potencial de acción (AP) más rápido genera un mejor seguimiento temporal de las entradas de alta frecuencia que conducen a una codificación y transferencia de información más eficiente en neuronas piramidales del lóbulo temporal. Reportan que “las neuronas piramidales corticales humanas de individuos con puntuaciones de CI más altas generan AP más rápidos” y por lo tanto “las neuronas de individuos con puntuaciones de CI más altas están mejor equipadas para procesar señales sinápticas a altas tasas y a escalas de tiempo más rápidas, lo cual es necesario para codificar grandes cantidades de información precisa y eficiente” (Goriounova, et al., 2018, p. e41724). Estos investigadores afirman que sus resultados ayudan a explicar por qué los tiempos de reacción más rápidos incluso en tareas simples (como medida de la velocidad de procesamiento mental) se asocian consistentemente con una mayor inteligencia general medida por puntajes de coeficiente intelectual en pruebas de inteligencia y otras medidas de g de Spearman (ver discusión de g, general factor inteligencia, en módulos anteriores de este capítulo).
También señalan que “las células piramidales son integradoras y acumuladoras de información sináptica. Las dendritas más grandes pueden contener físicamente más contactos sinápticos e integrar más información. De hecho, las dendritas de neuronas piramidales humanas reciben el doble de sinapsis que en roedores (DeFelipe et al., 2002) y la conectividad corticocortical de todo el cerebro se correlaciona positivamente con el tamaño de las dendritas en estas células” (Goriounova, et al., 2018, p. e41726). Goriounova et al. (2018) afirman que las matrices dendríticas más grandes y complejas en neuronas piramidales más grandes están acompañadas de aumentos en la integración en áreas corticales. “Las células piramidales humanas en las capas 2 y 3 [de la corteza cerebral] tienen dendritas 3 veces más grandes y más complejas que en macaco o ratón” que “sugieren una presión evolutiva tanto en la estructura dendrítica como en la forma de onda AP y enfatizan adaptaciones específicas de células piramidales humanas en áreas de asociación para cognitiva funciones” (p. e41727). Ellos razonan, por lo tanto, que las neuronas piramidales más grandes, con ramificaciones dendríticas más extensas y potenciales de inicio de acción más rápidos, son necesarias para el procesamiento cortical de orden superior asociado con la cognición humana. Con base en esta conclusión, sugieren que las diferencias entre individuos en “complejidad neuronal” podrían asociarse con diferencias entre personas en capacidad mental (Figura 14.8.5).
Figura\(\PageIndex{5}\): Una base celular de la inteligencia humana. Las puntuaciones de CI más altas se asocian con dendritas más grandes, potenciales de acción más rápidos durante la actividad neuronal y seguimiento de información más eficiente en neuronas piramidales de la corteza temporal. La cifra se basa en los resultados de Goriounova et al. (2018). (Imagen de Wikimedia Commons; Archivo:Base celular de IQ.png; https://commons.wikimedia.org/wiki/F...asis_of_IQ.png; por Natalia A. Goriounova y Huibert D. Mansvelder; licenciada bajo la licencia Creative Commons Attribution 4.0 International).
Resumen de Otras características cerebrales correlacionadas con diferencias en la inteligencia humana
En la siguiente tabla se resumen las características del cerebro asociadas a las diferencias individuales entre los humanos en la inteligencia general, medidas por el coeficiente intelectual y otras pruebas psicológicas de inteligencia. Basado en Goriounova y Mansvelder (2019).
====================================================================================================
Cuadro 14.5.1: Características neurológicas (filas no sombreadas) asociadas a la inteligencia (las filas 3 y 4 son continuaciones de las filas 1 y 2).
=====================================================================================================
| RASGO NEUROLÓGICO | volumen general del cerebro | Espesor cortical en corteza de asociación frontal, parietal y temporal | rápido aumento del grosor cortical en la infancia con disminución rápida del grosor cortical en la adolescencia temprana a medida que se produce la poda del exceso de sinapsis aumentando la eficiencia del procesamiento | eficiencia del procesamiento en áreas corticales distribuidas (frontal, parietal, temporal) medida por la menor actividad neural y la energía metabólica del cerebro inferior requerida para el desempeño de tareas mentales por individuos con alto coeficiente intelectual |
| CORRELACIÓN DEL TRATAMIENTO NEUROLÓGICO CON LA INTELIGENCIA MEDIANTE PRUEBAS PSICOLÓGICAS (r =, cuando lo declaran | positivo, r= entre .24 a .33 (correlacionado con el volumen cerebral general) |
correlaciones positivas con IQ (parte superior de esta columna anterior, es decir, espesor cortical) |
ambos cambios (aumento rápido seguido de disminución rápida) en el espesor cortical cortical se correlacionaron positivamente con un alto I.Q. | correlaciones positivas con inteligencia general “fluida” |
| Rasgo neurológico (continuación) | Estructura cortical y grosor cortical en áreas laterales de lóbulos temporales y en polo temporal | corteza prefrontal: estructura, función y conectividad | integridad funcional de la sustancia blanca (vías compuestas por axones mielinizados que interconectan regiones cerebrales), especialmente entre la corteza frontal derecha y temporal; retraso mental asociado con daño severo a la sustancia blanca | Volumen de materia gris del hemisferio izquierdo y conectividad de materia blanca (axones mielinizados) entre la corteza frontal orbital posterior izquierda (OFC) y la corteza cingulada anterior rostral (RACC) |
| CORRELACIÓN DEL TRATO NEUROLÓGICO CON LA INTELIGENCIA MEDIANTE | correlación positiva con inteligencia general “cristalizada” dependiente de habilidades verbales, memoria semántica de trabajo y conocimiento adquirido (correlacionada con la estructura y grosor cortical de lóbulos temporales y polo temporal) | correlación positiva especialmente con la capacidad de razonamiento (es decir, correlacionada con la estructura prefrontal, la función y la conectividad) | correlación positiva con inteligencia (es decir, correlacionada con la integridad de la materia blanca) | correlaciones positivas con inteligencia general “fluida”, “g” (ver sección 18.11) |
====================================================================================================
Resumen Conclusiones de la Tabla 14.5.1 (anterior): "Conclusiones sobre la distribución macroscópica cerebral de la inteligencia: la inteligencia está apoyada por una red distribuida de regiones cerebrales en muchas, si no todas, cortezas de asociación de orden superior, también conocidas como red parietal-frontal (Jung y Haier, 2007 ). Esta red incluye un gran número de regiones: la corteza prefrontal dorsolateral, el lóbulo parietal y el cingulado anterior, múltiples regiones dentro de los lóbulos temporal y occipital y, finalmente, tractos principales de materia blanca. Se puede observar alguna división limitada de la función, implicando áreas frontales y parietales en inteligencia fluida, lóbulos temporales en inteligencia cristalizada e integridad de la materia blanca en la velocidad de procesamiento. Aunque los estudios de imágenes cerebrales han identificado correlatos anatómicos y funcionales de la inteligencia humana, los coeficientes de correlación reales han sido consistentemente modestos, alrededor de 0.15—0.35" (Goriounova y Mansvelder, 2019). Obsérvese que la cantidad de variación (varianza) en una variable contabilizada por su correlación con otra variable es igual al coeficiente de correlación al cuadrado. Por ejemplo, en este caso, una correlación de 0.35 significa que solo .35 x .35 = .1225 (12.25%) de la variación en la inteligencia entre los individuos estudiados se atribuye a su correlación con las características anatómicas y funcionales de los cerebros estudiados en estudios de imágenes cerebrales, según Goriounova y Mansvelder ( 2019). Así, en el mejor de los casos, encuentran que alrededor del 88% de la varianza en la inteligencia entre los individuos estudiados se explica por las diferencias observadas entre individuos en las características anatómicas y funcionales de los cerebros de los estudiados mediante imágenes cerebrales. Quizás la imagen cerebral está detectando regiones cerebrales involucradas en solo algunos componentes de la inteligencia (¿quizás la atención?) mientras faltan otros como el procesamiento de similitud o las relaciones causales, por ejemplo.
Genes e Inteligencia
Los intentos de encontrar genes únicos que determinaran la inteligencia fracasaron por completo. La conclusión fue que la inteligencia debe ser un rasgo altamente poligénico. Sin embargo, la mayoría de los genes que afectan la inteligencia se encuentran en regiones no codificantes del ADN que no codifican la estructura proteica sino que son genes reguladores (que encienden y apagan otros genes) que están involucrados en la generación de neuronas corticales durante el desarrollo cerebral. Diferentes genes correlacionados con la inteligencia afectan principalmente los procesos prenatales de desarrollo cerebral incluyendo “la proliferación de células progenitoras neurales y su especialización, la migración de nuevas neuronas a las diferentes capas de la corteza, la proyección de axones de las neuronas a su diana de señalización y germinación dendrítica” (Goriounova y Mansvelder, 2019).
Los genes también afectan las interacciones célula-célula. “Muchos de los genes identificados que juegan un papel en el neurodesarrollo podrían contribuir a la función sináptica y plasticidad. La función cerebral se basa en procesos altamente dinámicos y dependientes de la actividad que encienden y apagan los genes. Estos pueden conducir a profundos cambios estructurales y funcionales e implican la formación de nuevas sinapsis y la eliminación de sinapsis no utilizadas, cambios en el citoesqueleto, la movilidad del receptor y el metabolismo energético. La capacidad cognitiva puede depender de cuán eficientes las neuronas puedan regular estos procesos... algunos genes candidatos.. están específicamente involucrados en la guía axónica durante el desarrollo neuronal” (Goriounova y Mansvelder, 2019). Esto puede sugerir que la eficiencia en las conexiones entre diferentes circuitos y áreas cerebrales puede ser un factor significativo en la inteligencia. Los axones que no van a los lugares correctos durante el desarrollo cerebral conducirían a ineficiencias de procesamiento.
Otros genes que pueden estar involucrados en diferencias entre individuos en inteligencia afectan las vías de señalización involucradas en la proliferación y migración de neuronas (durante el desarrollo cerebral prenatal; ver sección en el Capítulo 4 sobre el desarrollo del sistema nervioso) y la comunicación sináptica a lo largo del desarrollo.
Otros genes que pueden estar involucrados en las diferencias de inteligencia entre las personas son los genes que organizan actividades neuronales presinápticas, especialmente las vesículas sinápticas y su liberación de transmisor.
Otros regulan el cAMP y el CREB involucrados en la transcripción génica que para las neuronas juega un papel en la plasticidad sináptica, el aprendizaje y la memoria.
Otros juegan un papel en la función de los canales de calcio regulados por voltaje. Recordemos que los canales de calcio se abren cuando un potencial de acción alcanza un axón que termina estimulando el movimiento y la unión de vesículas sinápticas a la membrana axonal presináptica seguido de liberación del transmisor.
Curiosamente, la mayor parte de la energía consumida por el cerebro (20% del total del cuerpo) es para la generación de potenciales possinápticos (PSP). “Notablemente, la aparición de funciones cognitivas superiores en humanos durante la evolución también se asocia con el aumento de la expresión de genes del metabolismo energético” y “la capacidad cognitiva se asocia con la variación genética en varios genes que codifican para reguladores de la función mitocondrial” esenciales para el metabolismo energético .
“Además, los genes involucrados en el metabolismo lipídico (BTN2A1 y BTN1A1) y el metabolismo de la glucosa y aminoácidos (GPT). [así como] “proteínas asociadas a microtúbulos [que].. afectan el reciclaje de los receptores sinápticos y la liberación de neurotransmisores... vinculados a la inteligencia por varios estudios.. se encuentran entre los genes candidatos de la inteligencia” (Goriounova y Mansvelder, 2019). Se sabe que los genes que afectan a las proteínas asociadas a microtúbulos están alterados en la enfermedad de Alzheimer, Parkinson y Huntington.
La mayor expresión de los genes asociados a la inteligencia ocurre “dentro de las neuronas piramidales en el área del hipocampo CA1 y regiones somatosensoriales corticales. [y expresión significativa] en neuronas espinosas medias.. Las neuronas piramidales son los tipos neuronales más abundantes en neocórtex e hipocampo, estructuras asociadas con funciones ejecutivas superiores, toma de decisiones, resolución de problemas y memoria. Las neuronas espinosas medias estriatales constituyen el 95% de todos los tipos neuronales dentro del estriado, una estructura responsable de la motivación, la recompensa, el aprendizaje de hábitos y la producción conductual” (Goriounova y Mansvelder, 2019).
Un papel central para las neuronas piramidales e implicaciones de las comparaciones entre especies
Según Goriounova y Mansvelder (2019), “Los estudios genéticos indican que la expresión de genes asociados a la inteligencia se acumula en las neuronas piramidales corticales (Savage et al., 2018; Coleman et al., 2019). Las comparaciones de las propiedades celulares clave de las neuronas piramidales entre especies pueden ofrecer información sobre la significación funcional de tales diferencias para la cognición humana.. En comparación con roedores y macacos, las células piramidales de capa 2/3 humanas tienen dendritas tres veces más grandes y más complejas (Mohan et al., 2015 ). Además, estas dendrítas grandes también reciben dos veces más sinapsis que las neuronas piramidales de roedores (DeFelipe et al., 2002). Aparte de las diferencias estructurales, las neuronas piramidales humanas muestran una serie de propiedades funcionales únicas. las sinapsis excitatorias humanas se recuperan 3—4 veces más rápido de la depresión que las sinapsis en la corteza de roedores, tienen potenciales de acción más rápidos y transfieren información hasta nueve veces mayor que el ratón sinapsis (Testa-Silva et al., 2014).” En humanos, las neuronas piramidales más grandes, con dendritas más largas, ramificaciones dendríticas más complejas, conexiones más sinápticas y potenciales de inicio de acción más rápidos, se combinan para hacer posible el procesamiento de mayores cantidades de información, con mayor eficiencia e integración entre áreas cerebrales, que en otras animales, y estas neuronas grandes pueden desempeñar un papel central en las vastas diferencias entre la inteligencia humana y animal no humana (ver Figura 14.8.5).
Correlatos cerebrales de la creatividad
Jung y Haier (2013) reportan una serie de correlatos cerebrales de inteligencia y creatividad. Sin embargo, argumentan en contra de la idea de un área cerebral para una función cognitiva. En cambio, como se discutió en el módulo anterior, argumentan que las redes cerebrales que involucran múltiples áreas cerebrales están involucradas en la cognición, especialmente en procesos psicológicos complejos como la inteligencia y la creatividad. Sin embargo, reconocen que los estudios de lesiones y lesiones cerebrales revelan estructuras cerebrales que son necesarias, aunque no suficientes, para ciertas funciones psicológicas. Dan tres ejemplos: 1) Phineas Gage, quien sobrevivió a una varilla de hierro que pasaba por su lóbulo frontal resultando en cambios de personalidad y emocionales así como deterioro del juicio y pérdida de muchas inhibiciones sociales; 2) “Tan”, cuyo daño cerebral condujo a la identificación del área de Broca para expresión del lenguaje; y 3) H.M., cuya extirpación quirúrgica bilateral de estructuras de lóbulo temporal incluyendo hipocampo reveló el papel del hipocampo y estructuras relacionadas en la formación de nuevos recuerdos explícitos a largo plazo y su recuperación.
Dentro de este contexto, Jung y Haier (2013) señalan algunas observaciones interesantes a partir del examen post mortem del cerebro del famoso físico teórico, Albert Einstein (cuyo trabajo condujo a la ecuación, E=mC 2), y lo que podría sugerir sobre los mecanismos cerebrales en la creatividad. El cerebro de Einstein no era notable en muchos sentidos. Su tamaño y peso estuvieron dentro del rango normal para un hombre de su edad, y la morfología del lóbulo frontal y temporal y el área del cuerpo calloso no fueron diferentes de los cerebros control. Sin embargo, hubo una diferencia pronunciada. Según Jung y Haier, al cerebro de Einstein le faltaba el opérculo parietal, la localización típica de la corteza somatosensorial secundaria, resultando en un lóbulo parietal inferior más grande. En el cerebro de Einstein, el lóbulo parietal inferior era aproximadamente 15% más ancho que en los cerebros de los controles normales. Según Jung y Haier, esta región del cerebro está asociada con “la cognición visuoespacial, el razonamiento matemático y la imaginería del movimiento.. y su expansión se notó en otros casos de destacados físicos y matemáticos”. Agregan que un examen más detallado de esta área del cerebro de Einstein reveló que en lugar de más neuronas, esta región de su cerebro tenía un número mucho mayor de células gliales, que proporcionan nutrición a las neuronas, lo que quizás indica una cantidad inusualmente grande de actividad entre las neuronas en esta región de su cerebro.
Significativamente, como se describe en el módulo anterior, la corteza parietal tiene fuertes vínculos con la corteza prefrontal formando una red frontoparietal: el lóbulo parietal inferior es principalmente conectado con la corteza prefrontal dorsolateral (Bruner, 2010), asociado, en parte, con habilidades para el pensamiento abstracto, mientras que las regiones parietales superiores, según Bruner, como se discute en el módulo 14.2, se asocian en la literatura con funciones como representación abstracta, imágenes mentales internas, “mundo imaginado [s],.. y experimento de pensamiento” (es decir, imaginación). Jung y Haier detallan otro estudio de la corteza de asociación prefrontal derecha de Einstein, donde los investigadores encontraron una mayor densidad de empaquetamiento de neuronas (mismo número de neuronas en un espacio más pequeño), que se interpretó como más corta tiempos de conducción entre neuronas corticales en el cerebro de Einstein en comparación con cerebros de control. Jung y Haier concluyen que el cerebro de Einstein difería de los controles en la red frontoparietal. Estos autores han propuesto que la red frontoparietal es crucial para la inteligencia humana; además, plantearon la hipótesis de que las diferencias entre las personas en la eficiencia de la comunicación neuronal entre las regiones frontal y parietal de la corteza explica las diferencias en la inteligencia en humanos (Jung & Haier, 2007). En parte, esta idea se basa en su hallazgo de que las personas de alto coeficiente intelectual muestran menos actividad en estas áreas cerebrales durante una tarea cognitiva compleja, mientras que las personas con un coeficiente intelectual más bajo muestran más actividad cerebral, lo que sugiere que un coeficiente intelectual alto está relacionado con eficiencia en las operaciones de procesamiento de información neuronal. Además, el coeficiente intelectual más alto y la capacidad de abstracción se correlacionan inversamente con la tasa metabólica de la glucosa cerebral (Haier et al., 1988, 1992, 2003, 2004), lo que sugiere un modelo de eficiencia de diferencias individuales en g en el que aumenta la capacidad superior de abstracción eficiencia de procesamiento. En su Teoría de Integración Parietal-Frontal (P-FIT) de la base neural de la inteligencia, después del procesamiento sensorial, la información “se alimenta entonces hacia las cortezas parietales angulares, supramarginales e inferiores, donde el simbolismo estructural y/o abstracción se generan y manipulan. La corteza parietal interactúa entonces con regiones frontales que sirven para probar hipótesis diversas soluciones a un problema dado”. Agregan que “el cingulado anterior está involucrado en la selección de respuesta así como en la inhibición de respuestas competitivas. Este proceso depende críticamente de la fidelidad de la materia blanca subyacente necesaria para facilitar la transmisión rápida y libre de errores de datos entre lóbulos frontal y parietal "(Jung & Haier, 2013, p. 239). También señalan que la investigación en genética muestra que “la inteligencia y la estructura cerebral (es decir, materia gris y blanca) comparten genes comunes” (p. 240).
En cuanto a la creatividad específicamente, estos autores se refieren a una teoría de Flaherty (2005) que propone un sistema frontotemporal impulsado por la actividad límbica dopaminérgica que proporciona el impulso para la expresión creativa ya sea arte, música, escritura, ciencia, etc. y medido por pruebas de pensamiento divergente. Jung y Haier (2013) explican que el lóbulo temporal normalmente inhibe el lóbulo frontal de manera que la lesión o disfunción leve del lóbulo temporal libera actividad desde el lóbulo frontal por desinhibición provocando un aumento de las interacciones del lóbulo frontal con otras regiones cerebrales, a veces lo que lleva a un aumento de los resultados creativos de pacientes neurológicos con daño lateral izquierdo. Argumentan que este y otros datos de “tres estudios estructurales apuntan a una red decididamente lateralizada izquierda, frontosubcortical y desinhibitoria de regiones cerebrales subyacentes a la cognición creativa y al logro” (p. 244). Agregan que este modelo, que aún requiere mucha más investigación empírica, “parece incluir los lóbulos frontal y temporal, modulándose el “tono” cortical a través de interacciones entre los lóbulos frontales, los ganglios basales y el tálamo (parte del sistema de dopamina) a través de vías de materia blanca” (p. 244). Aunque este modelo es especulativo para una forma tan compleja de cognición como la creatividad, puede guiar la investigación continua sobre cómo los humanos desarrollan productos creativos intelectuales y artísticos.
| Lóbulo parietal inferior | |
|---|---|
|
Figura\(\PageIndex{6}\): Superficie lateral del hemisferio cerebral izquierdo. El lóbulo parietal inferior se muestra en naranja. (Imagen de Wikimedia Commons; Archivo:Gray726 parietal inferior lobule.png; https://commons.wikimedia.org/wiki/F...tal_lobule.png; por Gray, vectorizado por Mysid, colourd por was_a_bee.; esta obra es de dominio público. Esto se aplica en todo el mundo). |
|
|
Figura\(\PageIndex{7}\): Anatomía superficial del lóbulo parietal inferior. Morado: Circunvolución supramarginal. Azul: Circulación angular. LS: Surco lateral (fisura silviana). CS: Surco central. IPS: Surco intraparietal. STS:Surco temporal superior. PN: Muesca preoccipital. (Imagen y subtítulo de Wikimedia Commons; Archivo:Anatomía superficial del lóbulo parietal inferior (IPL) .png); https://commons.wikimedia.org/wiki/F...bule_(IPL).png; por Joshua D. Burks Lillian B. Boettcher Andrew K. Conner Chad A. Glenn Phillip A. Bonney Cordell M. Baker Robert G. Briggs Nathan A. Pittman Daniel L. O'Donoghue Dee H. Wu Michael E. Sughrue; licenciado bajo la licencia Creative Commons Attribution 4.0 International). |
.png)
Pruebas de Inteligencia y Concepciones de Inteligencia Humana
El desarrollo de pruebas para medir la inteligencia ha tenido un gran impacto en el desarrollo de ideas sobre la naturaleza y estructura de la inteligencia humana, y su base biológica en el cerebro. La mayoría de las teorías de la inteligencia humana se basan en datos derivados de pruebas de inteligencia, datos que se analizan mediante análisis factorial, un método matemático para analizar patrones de correlaciones entre diferentes medidas de habilidades mentales. En la sección 14.2, ya hemos discutido cómo este método, inventado y utilizado por Spearman (1904), reveló el factor “g” en la inteligencia humana.
Para comprender el pensamiento actual y la investigación sobre las bases biológicas de la inteligencia humana, es esencial adquirir al menos una familiaridad general con los principales modelos teóricos de la inteligencia humana que los psicólogos han desarrollado. Para ello, este capítulo cuenta con una sección complementaria titulada “Modelos Tradicionales de Inteligencia Humana”. Es muy recomendable que los alumnos lean esa sección complementaria. Las teorías que examinamos en esa sección se basan en gran medida en pruebas de inteligencia y análisis factorial, mientras que otras son más intuitivas. Esa sección también introduce figuras históricas clave, grandes teorías de la inteligencia y estrategias comunes de evaluación utilizadas para medir la inteligencia humana.
En la sección 14.2, discutimos una serie de regularidades universales del entorno perdurables, generacionales, que han sido incorporadas por la evolución en la organización cerebral y la inteligencia. Como se describe en esa sección, estos hechos perdurables sobre cómo funciona el mundo incluyen información innata, genéticamente internalizada sobre objetos en el espacio tridimensional, el paso del tiempo, ciclos diarios de luz y oscuridad, relaciones de causalidad (formando base para la lógica causal e inferencia), relaciones de similitud (que conducen a la formación de categorías y lógica e inferencia categóricas), y las relaciones predictivas, basadas en la covariación de eventos, permitiendo que los cerebros humanos y animales proyecten mentalmente el organismo hacia el futuro. Todas estas propiedades invariantes del mundo deben incluirse en los modelos neuronales del cerebro o mapas cognitivos del mundo para que el cerebro guíe eficazmente el comportamiento adaptativo.
Cuando examinemos los modelos tradicionales de inteligencia en la sección suplementaria antes mencionada, reconocerá que cada uno se enfoca en una sola, o algunas de las facetas de la inteligencia discutidas en el enfoque evolutivo tomado en la sección 14.2. En cierto sentido, cada teoría discutida en esa sección es similar a la fábula de los ciegos que intentan describir a un elefante. Cada ciego sólo conoce esa parte del elefante que pasa a sentir y así cada hombre tiene una comprensión diferente e incompleta del conjunto. De igual manera, cada teoría de la inteligencia se enfoca solo en parte del complejo de procesos a los que colectivamente denominamos “inteligencia”. Sin embargo, cada teoría hace una contribución, y cada una, de una o más maneras, se relaciona con la discusión evolutiva en la sección 14.2.
Por ejemplo, como verás, la inteligencia emocional, incluida la inteligencia intra e interpersonal de Gardner, está relacionada con representaciones neuronales de las contingencias del entorno social, mecanismos cerebrales para los cuales son el foco del nuevo campo de la neurociencia cognitiva social. Las inteligencias múltiples de Gardner incluyen inteligencia espacial relacionada con la representación de objetos en el espacio tridimensional, habilidades que requieren porciones de corteza parietal e hipocampo. En el nivel III en la teoría de la inteligencia de Carroll se encuentra “g”, la inteligencia general, relacionada con representaciones de relaciones causales, de similitud y predictivas, probablemente involucrando la red frontoparietal (Jung & Haier, 2007). De estas teorías, la primera, la teoría de tres estratos de la inteligencia humana de Carroll es, con mucho, la más aceptada y productiva en términos de poder explicativo y evidencia empírica. Con estos antecedentes, estarás mejor preparado para entender los modelos tradicionales de inteligencia y, quizás lo más importante para este curso, obtendrás una mejor comprensión de las bases biológicas de la inteligencia y el pensamiento, el foco principal de este capítulo.
Referencias
Assem, Moataz, Blank, Idan A, Mineroff, Zachary, Ademoğlu, Ahmet y Fedorenko, Evelina. (2020). “La actividad en la red fronto-parietal de múltiples demandas está fuertemente asociada con diferencias individuales en la memoria de trabajo y la inteligencia fluida”. Cortex, 131.
Brissenden, J. A., Levin, E. J., Osher, D. E., Halko, M. A., & Somers, D. C. (2016). Evidencia funcional de un nódulo cerebeloso de la red de atención dorsal. Revista de Neurociencia, 36 (22), 6083-6096.
Dixon, M. L., Andrews-Hanna, J. R., Spreng, R. N., Irving, Z. C., Mills, C., Girn, M., & Christoff, K. (2017). Las interacciones entre la red predeterminada y la red de atención dorsal varían según los subsistemas predeterminados, el tiempo y los estados cognitivos. Neuroimagen, 147, 632-649.
Duncan, J. (2010). El sistema de demanda múltiple (MD) del cerebro de primates: programas mentales para el comportamiento inteligente. Tendencias en ciencias cognitivas, 14 (4), 172-179.
Duncan, J. (2013). La estructura de la cognición: episodios atencionales en la mente y el cerebro. Neurona 80:35 —50. Disponible en: http://dx.doi.org/10.1016/j.neuron.2013.09.015
Goriounova, N. A., Heyer, D. B., Wilbers, R., Verhoog, M. B., Giugliano, M., Verbist, C.,... & Mansvelder, H. D. (2018). Las neuronas piramidales humanas grandes y rápidas se asocian con la inteligencia. Elife, 7, e41714.
Goriounova, N. A., & Mansvelder, H. D. (2019). Genes, células y áreas cerebrales de inteligencia. Fronteras en la neurociencia humana, 44.
Gray, J. R., Chabris, C. F., & Braver, T. S. (2003). Mecanismos neuronales de la inteligencia fluida general. Neurociencia de la naturaleza, 6 (3), 316-322.
Haier, R. J., Siegel Jr, B. V., Nuechterlein, K. H., Hazlett, E., Wu, J. C., Paek, J., Browning, H.L. y Buchsbaum, M. S. (1988). La tasa metabólica de glucosa cortical correlaciona el razonamiento abstracto y la atención estudiada con tomografía por emisión de positrones Inteligencia, 1 2 (2) ,199-217.
Haier, R. J., Siegel, B., Tang, C., Abel, L., y Buchsbaum, M. S. (1992). Inteligencia y cambios en la tasa metabólica regional de la glucosa cerebral después del aprendizaje. Inteligencia 16 (3), 415.
Haier, R. J., White, N. S., y Alkire, M. T. (2003). Las diferencias individuales en la inteligencia general se correlacionan con la función cerebral durante tareas no razonadas. Inteligencia, 31 (5), 429-441.
Haier R.J, Jung R., Yeo R., Jefe K., Alkire M.T. (2004). Variación cerebral estructural e inteligencia general. NeuroImage, 23 (1), 425-433.
Hearne, L. J., Mattingley, J. B., & Cocchi, L. (2016). Redes cerebrales funcionales relacionadas con diferencias individuales en inteligencia humana en reposo. Informes científicos, 6 (1), 1-8.
Jung, R. E., & Haier, R. J. (2007). La Teoría de Integración Parieto-Frontal (P-FIT) de la inteligencia: evidencia convergente de neuroimagen. Ciencias del comportamiento y del cerebro, 30 (2), 135-154.
Jung, R. E., & Haier, R. J. (2013). Creatividad e inteligencia: redes cerebrales que vinculan y diferencian la expresión del genio
Koenigshofer, K. A. (2017). Inteligencia general: adaptación a invariantes relacionales abstractas evolutivamente familiares, no a la novedad ambiental o evolutiva. El diario de la mente y el comportamiento, 38 (2), 119-153.
Rueda, M. R., Pozuelos, J. P., & Cómbita, L. M. (2015). Neurociencia cognitiva de la atención desde los mecanismos cerebrales hasta las diferencias individuales en la eficiencia. AIMS Neurociencia, 2 (4), 183-202.
Salinas E. & Sejnowski TJ. (2001). La actividad neuronal correlacionada y el flujo de información neuronal. Nature Reseñas Neurociencia 2:539—550. DOI: https://doi.org/10.1038/35086012, PMID: 11483997
Spearman, C. (1904). “Inteligencia General”, Objetivamente Determinada y Medida. The American Journal of Psychology, 15 (2), 201-292.
Atribuciones
“Mecanismos cerebrales e inteligencia”, escrito por Kenneth A. Koenigshofer, Ph.D., está licenciado bajo CC BY 4.0.