
2.4: Hormonas y Comportamiento
- Page ID
- 146506

La Universidad Estatal de Ohio
El objetivo de este módulo es introducirte en el tema de las hormonas y el comportamiento. Este campo de estudio también se denomina endocrinología conductual, que es el estudio científico de la interacción entre las hormonas y el comportamiento. Esta interacción es bidireccional: las hormonas pueden influir en el comportamiento, y el comportamiento a veces puede influir en las concentraciones hormonales. Las hormonas son mensajeros químicos liberados de las glándulas endocrinas que viajan a través del sistema sanguíneo para influir en el sistema nervioso para regular comportamientos como la agresión, el apareamiento y la crianza de los individuos.
objetivos de aprendizaje
- Definir la terminología básica y los principios básicos de las interacciones hormona-comportamiento.
- Explicar el papel de las hormonas en la diferenciación sexual conductual.
- Explicar el papel de las hormonas en el comportamiento agresivo.
- Explicar el papel de las hormonas en el comportamiento parental.
- Proporcione ejemplos de algunas interacciones comunes entre la hormona y el comportamiento.
Introducción
Este módulo describe la relación entre las hormonas y el comportamiento. Es probable que muchos lectores ya estén familiarizados con la idea general de que las hormonas pueden afectar el comportamiento. Los estudiantes generalmente están familiarizados con la idea de que las concentraciones de hormonas sexuales aumentan en la sangre durante la pubertad y disminuyen a medida que envejecemos, especialmente después de aproximadamente 50 años de edad. El comportamiento sexual muestra un patrón similar. La mayoría de las personas también conocen la relación entre la agresión y las hormonas esteroides anabólicas, y saben que la administración de hormonas esteroides artificiales a veces resulta en un comportamiento incontrolable y violento llamado “furia roid”. Muchas hormonas diferentes pueden influir en varios tipos de comportamiento, pero para el propósito de este módulo, limitaremos nuestra discusión a solo algunos ejemplos de hormonas y comportamientos. Por ejemplo, ¿las diferencias de sexo conductual son el resultado de las hormonas, el ambiente o alguna combinación de factores? ¿Por qué los hombres son mucho más propensos que las mujeres a cometer actos agresivos? ¿Están las hormonas involucradas en mediar el llamado “instinto” materno? Los endocrinólogos conductuales están interesados en cómo los efectos fisiológicos generales de las hormonas alteran el desarrollo y la expresión del comportamiento y cómo el comportamiento puede influir en los efectos de las hormonas. Este módulo describe, tanto fenomenológica como funcionalmente, cómo las hormonas afectan el comportamiento.
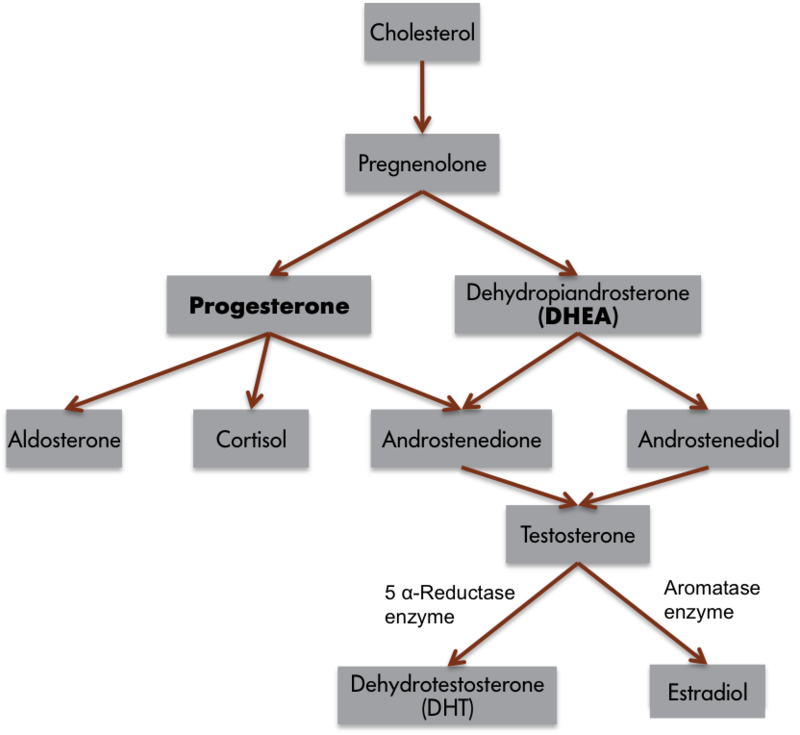
Para entender la relación hormona-comportamiento, es importante describir brevemente las hormonas. Las hormonas son mensajeros químicos orgánicos producidos y liberados por glándulas especializadas llamadas glándulas endocrinas. Las hormonas se liberan de estas glándulas a la sangre, donde pueden viajar para actuar sobre las estructuras diana a cierta distancia de su origen. Las hormonas son similares en función a los neurotransmisores, los químicos utilizados por el sistema nervioso en la coordinación de las actividades de los animales. Sin embargo, las hormonas pueden operar a mayor distancia y en un rango temporal mucho mayor que los neurotransmisores (Focus Topic 1). Ejemplos de hormonas que influyen en el comportamiento incluyen hormonas esteroides como la testosterona (un tipo común de andrógeno), estradiol (un tipo común de estrógeno), progesterona (un tipo común de progestina) y cortisol (un tipo común de glucocorticoide) (Tabla 1, A-B). Varios tipos de hormonas proteicas o peptídicas (pequeñas proteínas) también influyen en el comportamiento, incluyendo oxitocina, vasopresina, prolactina y leptina.
Tema de Enfoque 1: Transmisión Neural versus Comunicación Hormonal
Aunque la comunicación neuronal y hormonal se basan en señales químicas, existen varias diferencias prominentes. La comunicación en el sistema nervioso es análoga a viajar en un tren. Puedes usar el tren en tus planes de viaje siempre y cuando existan vías entre tu origen y destino propuesto. Asimismo, los mensajes neuronales pueden viajar solo a destinos a lo largo de los tractos nerviosos existentes. La comunicación hormonal, por otro lado, es como viajar en un automóvil. Se puede conducir a muchos más destinos de lo que permite el viaje en tren porque hay muchas más carreteras que vías del ferrocarril. De igual manera, los mensajes hormonales pueden viajar a cualquier parte del cuerpo a través del sistema circulatorio; cualquier célula que reciba sangre es potencialmente capaz de recibir un mensaje hormonal.
La comunicación neuronal y hormonal también difieren en otras formas. Para ilustrarlos, considere las diferencias entre las tecnologías digitales y analógicas. Los mensajes neuronales son eventos digitales, todo o ninguno que tienen un rápido inicio y desplazamiento: las señales neuronales pueden tener lugar en milisegundos. En consecuencia, el sistema nervioso media cambios en el cuerpo que son relativamente rápidos. Por ejemplo, el sistema nervioso regula la ingesta inmediata de alimentos y dirige el movimiento corporal. Por el contrario, los mensajes hormonales son eventos analógicos y graduados que pueden tardar segundos, minutos o incluso horas en ocurrir. Las hormonas pueden mediar procesos a largo plazo, como el crecimiento, el desarrollo, la reproducción y el metabolismo.
Los mensajes hormonales y neuronales son de naturaleza química, y son liberados y recibidos por las células de manera similar; sin embargo, también hay diferencias importantes. Los neurotransmisores, los mensajeros químicos utilizados por las neuronas, viajan una distancia de solo 20—30 nanómetros (30 X 10—9 m) —hasta la membrana de la neurona postsináptica, donde se unen con receptores. Las hormonas ingresan al sistema circulatorio y pueden viajar de 1 milímetro a >2 metros antes de llegar a una célula diana, donde se unen con receptores específicos.
Otra distinción entre comunicación neural y hormonal es el grado de control voluntario que se puede ejercer sobre su funcionamiento. En general, hay más control voluntario de señales neuronales que hormonales. Es prácticamente imposible lograr un cambio en tus niveles de hormona tiroidea, por ejemplo, mientras que mover tus extremidades al mando es fácil.
Si bien se trata de diferencias significativas, la división entre el sistema nervioso y el sistema endocrino se vuelve más borrosa a medida que aprendemos más sobre cómo el sistema nervioso regula la comunicación hormonal. Una mejor comprensión de la interfaz entre el sistema endocrino y el sistema nervioso, llamada neuroendocrinología, es probable que produzca avances importantes en el estudio futuro de la interacción entre hormonas y comportamiento.


Las hormonas coordinan la fisiología y el comportamiento de los individuos regulando, integrando y controlando las funciones corporales. Durante el tiempo evolutivo, las hormonas a menudo han sido cooptadas por el sistema nervioso para influir en el comportamiento para asegurar el éxito reproductivo. Por ejemplo, las mismas hormonas, la testosterona y el estradiol, que provocan la maduración del gameto (óvulo o esperma) también promueven el comportamiento de apareamiento. Esta doble función hormonal asegura que el comportamiento de apareamiento ocurre cuando los animales tienen gametos maduros disponibles para la fertilización. Otro ejemplo de regulación endocrina de la función fisiológica y conductual lo proporciona el embarazo. Las concentraciones de estrógenos y progesterona son elevadas durante el embarazo, y estas hormonas suelen estar involucradas en la mediación del comportamiento materno en las madres.
No todas las células están influenciadas por todas y cada una de las hormonas. Más bien, cualquier hormona dada puede influir directamente solo en las células que tienen receptores hormonales específicos para esa hormona en particular. Las células que tienen estos receptores específicos se denominan células diana para la hormona. La interacción de una hormona con su receptor inicia una serie de eventos celulares que eventualmente conducen a la activación de vías enzimáticas o, alternativamente, enciende o apaga la activación génica que regula la síntesis de proteínas. Las proteínas recién sintetizadas pueden activar o desactivar otros genes, causando otra cascada de eventos celulares. Es importante destacar que debe estar disponible un número suficiente de receptores hormonales apropiados para que una hormona específica produzca cualquier efecto. Por ejemplo, la testosterona es importante para el comportamiento sexual masculino. Si los hombres tienen muy poca testosterona, entonces la motivación sexual puede ser baja y puede restaurarse mediante el tratamiento con testosterona. Sin embargo, si los hombres tienen niveles normales o incluso elevados de testosterona pero muestran un bajo impulso sexual, entonces podría ser posible que la falta de receptores sea la causa y el tratamiento con hormonas adicionales no será efectivo.
¿Cómo podrían afectar las hormonas al comportamiento? En términos de su comportamiento, se puede pensar en humanos y otros animales conceptualmente como compuestos por tres componentes que interactúan: (1) sistemas de entrada (sistemas sensoriales), (2) integradores (el sistema nervioso central) y (3) sistemas de salida, o efectores (por ejemplo, músculos). Las hormonas no provocan cambios de comportamiento. Más bien, las hormonas influyen en estos tres sistemas de manera que los estímulos específicos son más propensos a provocar ciertas respuestas en el contexto conductual o social apropiado. Es decir, las hormonas cambian la probabilidad de que se emita un comportamiento particular en la situación apropiada (Nelson, 2011). Esta es una distinción crítica que puede afectar la forma en que pensamos de las relaciones hormona-comportamiento.
Podemos aplicar este esquema conductual de tres componentes a un comportamiento simple, cantando en pinzones cebra. Sólo cantan pinzones cebra machos. Si se extraen los testículos de pinzones machos adultos, entonces las aves reducen el canto, pero los pinzones castrados vuelven a cantar si los testículos son reimplantados, o si las aves son tratadas con testosterona o estradiol. Aunque comúnmente consideramos que los andrógenos son hormonas “masculinas” y los estrógenos como hormonas “femeninas”, es común que la testosterona se convierta en estradiol en las células nerviosas (Figura 1.5.1). Por lo tanto, ¡muchos comportamientos masculinos están asociados con las acciones de los estrógenos! De hecho, todos los estrógenos deben convertirse primero a partir de andrógenos debido al típico proceso de síntesis bioquímica. Si la enzima convertidora es baja o falta, entonces es posible que las hembras produzcan andrógenos excesivos y posteriormente desarrollen rasgos masculinos asociados. También es posible que los estrógenos en el ambiente afecten el sistema nervioso de los animales, incluidas las personas (por ejemplo, Kidd et al., 2007). Nuevamente, el comportamiento de canto es más frecuente cuando las concentraciones de testosterona o estrógeno en sangre son altas. Los machos cantan para atraer parejas o alejar a posibles competidores de sus territorios.
Si bien es evidente a partir de estas observaciones que los estrógenos están involucrados de alguna manera en el canto, ¿cómo podría el marco de tres componentes que se acaba de introducir ayudarnos a formular hipótesis para explorar el papel del estrógeno en este comportamiento? Al examinar los sistemas de entrada, podríamos determinar si los estrógenos alteran las capacidades sensoriales de las aves, haciendo más sobresalientes las señales ambientales que normalmente provocan el canto. Si este fuera el caso, entonces las mujeres o competidores podrían verse o escucharse más fácilmente. Los estrógenos también podrían influir en el sistema nervioso central. La arquitectura neuronal o la velocidad del procesamiento neural podrían cambiar en presencia de estrógenos. Los procesos neuronales superiores (por ejemplo, motivación, atención o percepción) también podrían verse influenciados. Finalmente, los órganos efectores, los músculos en este caso, podrían verse afectados por la presencia de estrógenos. Las concentraciones de estrógeno en la sangre podrían afectar de alguna manera los músculos de la siringe de un pájaro cantor (el órgano vocal de las aves). Los estrógenos, por lo tanto, podrían afectar el canto de los pájaros al influir en las capacidades sensoriales, el sistema central de procesamiento u órganos efectores de un ave individual. No entendemos completamente cómo el estrógeno, derivado de la testosterona, influye en el canto de los pájaros, pero en la mayoría de los casos, se puede considerar que las hormonas afectan el comportamiento al influir en uno, dos o los tres componentes, y este marco de tres partes puede ayudar en el diseño de hipótesis y experimentos para explorar estos temas.
¿Cómo podrían afectar las conductas a las hormonas? El ejemplo del canto de los pájaros demuestra cómo las hormonas pueden afectar el comportamiento, pero como se señaló, también se produce la relación recíproca; es decir, el comportamiento puede afectar las concentraciones hormonales. Por ejemplo, la visión de un intruso territorial puede elevar las concentraciones de testosterona en sangre en aves macho residentes y así estimular el comportamiento de canto o lucha. De igual manera, los ratones machos o monos rhesus que pierden una pelea disminuyen las concentraciones circulantes de testosterona durante varios días o incluso semanas después. También se han reportado resultados comparables en humanos. Las concentraciones de testosterona se ven afectadas no sólo en humanos involucrados en combate físico, sino también en aquellos involucrados en batallas simuladas. Por ejemplo, las concentraciones de testosterona se elevaron en los ganadores y disminuyeron en los perdedores de los torneos regionales de ajedrez.

Las personas no tienen que involucrarse directamente en un concurso para que sus hormonas se vean afectadas por el resultado del concurso. Se reclutó a fanáticos masculinos tanto de la selección brasileña como de la italiana para proporcionar muestras de saliva para ser analizadas en busca de testosterona antes y después del juego final del partido de fútbol de la Copa del Mundo en 1994. Brasil e Italia estaban empatados al entrar en el juego final, pero Brasil ganó con un tiro penal en el último momento posible. La afición brasileña estaba eufórica y la afición italiana quedó coronada. Cuando se ensayaron las muestras, 11 de los 12 fanáticos brasileños que fueron muestreados tuvieron mayores concentraciones de testosterona, y 9 de 9 fans italianos tenían concentraciones de testosterona disminuidas, en comparación con los valores basales previos al juego (Dabbs, 2000).
En algunos casos, las hormonas pueden verse afectadas por la anticipación del comportamiento. Por ejemplo, las concentraciones de testosterona también influyen en la motivación sexual y el comportamiento en las mujeres. En un estudio, la interacción entre las relaciones sexuales y la testosterona se comparó con otras actividades (caricias o ejercicio) en mujeres (van Anders, Hamilton, Schmidt, & Watson, 2007). En tres ocasiones distintas, las mujeres proporcionaron una muestra de pre-actividad, post-actividad y saliva a la mañana siguiente. Después del análisis, se determinó que la testosterona de las mujeres estaba elevada antes del coito en comparación con otras ocasiones. Así, existe una relación anticipatoria entre la conducta sexual y la testosterona. Los valores de testosterona fueron más altos después de la relación sexual en comparación con el ejercicio, lo que sugiere que participar en el comportamiento sexual también puede influir en las concentraciones hormonales
Diferencias de sexo
Las gallinas y gallos son diferentes. Las vacas y los toros son diferentes. Hombres y mujeres son diferentes. Incluso las niñas y los niños son diferentes. Los humanos, como muchos animales, son sexualmente dimórficos (di, “dos”; morph, “type”) en el tamaño y la forma de sus cuerpos, su fisiología, y para nuestros propósitos, su comportamiento. El comportamiento de niños y niñas difiere en muchos sentidos. Las niñas generalmente sobresalen en habilidades verbales en relación con los niños; los niños tienen casi el doble de probabilidades que las niñas de sufrir dislexia (dificultades de lectura) y tartamudeo y casi 4 veces más probabilidades de sufrir autismo. Los niños son generalmente mejores que las niñas en tareas que requieren habilidades visuoespaciales. Las niñas se involucran en comportamientos de crianza con más frecuencia que los niños. Más del 90% de todos los casos de anorexia nerviosa involucran a mujeres jóvenes. Los hombres jóvenes tienen el doble de probabilidades que las mujeres jóvenes de sufrir esquizofrenia. Los niños son mucho más agresivos y generalmente participan en juegos más rudos que las niñas (Berenbaum, Martin, Hanish, Briggs, & Fabes, 2008). Muchas diferencias de sexo, como la diferencia de agresividad, persisten a lo largo de la edad adulta. Por ejemplo, hay muchos más hombres que mujeres que cumplen penas de prisión por comportamiento violento. Las diferencias hormonales entre hombres y mujeres pueden explicar las diferencias de sexo adulto que se desarrollan durante la pubertad, pero ¿qué explica las diferencias de sexo conductual entre los niños antes de la pubertad y la activación de sus gónadas? Las secreciones hormonales de las gónadas en desarrollo determinan si el individuo se desarrolla de manera masculina o femenina. Los testículos embrionarios de mamíferos producen andrógenos, así como hormonas peptídicas, que dirigen el desarrollo del cuerpo, el sistema nervioso central y el comportamiento posterior en dirección masculina. Los ovarios embrionarios de los mamíferos son prácticamente inactivos y no secretan altas concentraciones de hormonas. En presencia de ovarios, o en ausencia total de gónadas, el desarrollo morfológico, neural y, posteriormente, conductual sigue una vía femenina.

Las hormonas esteroides gonadales tienen efectos organizativos (o de programación) sobre el cerebro y el comportamiento (Phoenix, Goy, Gerall, & Young, 1959). Los efectos organizativos de las hormonas esteroides están relativamente limitados a las primeras etapas de desarrollo. Existe una asimetría en los efectos de los testículos y ovarios sobre la organización del comportamiento en mamíferos. La exposición hormonal temprana en la vida tiene efectos organizativos sobre el comportamiento posterior de los roedores; el tratamiento temprano con hormonas esteroideas provoca una masculinización relativamente irreversible y permanente del comportamiento de los roedores (apareamiento y agresivo). Estos efectos hormonales tempranos se pueden contrastar con las influencias conductuales reversibles de las hormonas esteroides proporcionadas en la edad adulta, las cuales se denominan efectos activacionales. Los efectos activacionales de las hormonas sobre el comportamiento de los adultos son temporales y pueden disminuir poco después de metabolizar la hormona. Así, el comportamiento masculino típico requiere la exposición a andrógenos durante la gestación (en humanos) o inmediatamente después del nacimiento (en roedores) para masculinizar algo el cerebro y también requiere andrógenos durante o después de la pubertad para activar estos circuitos neuronales. El comportamiento típico femenino requiere una falta de exposición a los andrógenos temprano en la vida lo que conduce a la feminización del cerebro y también requiere de estrógenos para activar estos circuitos neuronales en la edad adulta. Pero esta simple dicotomía, que funciona bien con animales con dimorfismo sexual muy distinto en el comportamiento, tiene muchas advertencias cuando se aplica a las personas.
Si caminas por alguna juguetería importante, entonces probablemente observarás un par de pasillos llenos de cajas rosadas y la ausencia total de empaques rosados de juguetes en pasillos adyacentes. Sorprendentemente, también verás una fuerte autosegregación de niños y niñas en estos pasillos. Es raro ver chicos en los pasillos “rosados” y viceversa. A los fabricantes de juguetes a menudo se les acusa de fabricar juguetes que tienen prejuicios de género, pero parece más probable que niños y niñas disfruten jugando con tipos y colores específicos de juguetes. En efecto, los fabricantes de juguetes duplicarían inmediatamente sus ventas si pudieran vender juguetes a ambos sexos. Los niños generalmente prefieren juguetes como camiones y pelotas y las niñas generalmente prefieren juguetes como muñecas. Si bien es dudoso que existan genes que codifican preferencias para los coches de juguete y camiones en el cromosoma Y, es posible que las hormonas puedan dar forma al desarrollo del cerebro de un niño para preferir ciertos tipos de juguetes o estilos de comportamiento de juego. Es razonable creer que los niños aprenden qué tipos de juguetes y qué estilos de juego son apropiados a su género. ¿Cómo podemos entender y separar la contribución de los mecanismos fisiológicos del aprendizaje a entender las diferencias de sexo en los comportamientos humanos? Para desenredar estos temas, a menudo se utilizan modelos animales. A diferencia de la situación en los humanos, donde las diferencias de sexo suelen ser solo una cuestión de grado (a menudo leve), en algunos animales, los miembros de un solo sexo pueden mostrar un comportamiento particular. Como se señaló, a menudo solo cantan los pájaros cantores machos. Los estudios de tales comportamientos fuertemente sesgados por el sexo son particularmente valiosos para comprender la interacción entre el comportamiento, las hormonas y el sistema nervioso.
Un estudio de monos vervet pone en duda la primacía del aprendizaje en el establecimiento de preferencias de juguete (Alexander & Hines, 2002). Los monos vervet hembra preferían los juguetes típicos de las niñas, como muñecas o ollas para cocinar, mientras que los monos vervet machos preferían los juguetes típicos de los niños, como autos o pelotas. No hubo diferencias de sexo en la preferencia por los juguetes neutrales de género, como libros ilustrados o peluches. Presumiblemente, los monos no tienen un concepto previo de juguetes de “niño” o “niña”. Los monos rhesus jóvenes también muestran preferencias de juguete similares.

¿Qué subyace entonces a la diferencia de sexo en la preferencia por juguetes? Es posible que ciertos atributos de los juguetes (u objetos) atraigan ya sea a niños o niñas. Los juguetes que atraen a los niños o a los monos vervet o rhesus masculinos, en este caso, una pelota o un carro de juguete, son objetos que se pueden mover activamente por el espacio, juguetes que se pueden incorporar en el juego activo, áspero y de caída. El atractivo de los juguetes que prefieren las niñas o las hembras de monos vervet parece estar basado en el color. El rosa y el rojo (los colores de la muñeca y la olla) pueden provocar la atención a los bebés.
La sociedad puede reforzar esas respuestas estereotipadas a los juguetes típicos de género. Las diferencias de sexo en las preferencias de juguetes surgen a los 12 o 24 meses de edad y parecen fijas a los 36 meses de edad, pero ¿las diferencias de sexo en la preferencia de juguete están presentes durante el primer año de vida? Es difícil preguntar a los bebés preverbales qué prefieren, pero en estudios donde los investigadores examinaron la cantidad de tiempo que los bebés miraron diferentes juguetes, los datos de seguimiento ocular indican que bebés de tan solo 3 meses mostraron diferencias de sexo en las preferencias de juguetes; las niñas preferían muñecas, mientras que los niños prefirieron camiones. Otro resultado que sugiere, pero no prueba, que las hormonas están involucradas en las preferencias de los juguetes es la observación de que las niñas diagnosticadas con hiperplasia suprarrenal congénita (CAH), cuyas glándulas suprarrenales producen cantidades variables de andrógenos temprano en la vida, jugaban con juguetes masculinos con mayor frecuencia que las niñas sin CAH. Además, se observó una relación dosis-respuesta entre la extensión del trastorno (es decir, el grado de exposición a andrógenos fetales) y el grado de masculinización del comportamiento lúdico. ¿Son las diferencias de sexo en las preferencias de juguete o la actividad lúdica, por ejemplo, las consecuencias inevitables de los entornos endocrinos diferenciales de niños y niñas, o estas diferencias son impuestas por prácticas y creencias culturales? ¿Estas diferencias son el resultado de recibir juguetes específicos de género desde una edad temprana, o son estas diferencias alguna combinación de factores endocrinos y culturales? Nuevamente, estas son preguntas difíciles de desentrañar en las personas.
Incluso cuando las diferencias de sexo conductual aparecen temprano en el desarrollo, parece haber alguna duda con respecto a las influencias de las expectativas sociales. Un ejemplo es el patrón de comportamiento de juego humano durante el cual los machos son más físicos; este patrón se ve en otras especies, incluyendo primates no humanos, ratas y perros. ¿La diferencia en la frecuencia de juego rudo entre niños y niñas se debe a factores biológicos asociados con ser masculino o femenino, o se debe a expectativas culturales y aprendizaje? Si hay una combinación de influencias biológicas y culturales que median la frecuencia del juego rudo y caída, entonces ¿qué proporción de la variación entre los sexos se debe a factores biológicos y qué proporción se debe a las influencias sociales? Es importante destacar, ¿es apropiado hablar de diferencias de sexo “normales” cuando estos rasgos prácticamente siempre se organizan a lo largo de un continuo en lugar de en categorías discretas?
Las diferencias de sexo son comunes en humanos y en animales no humanos. Debido a que hombres y mujeres difieren en la proporción de concentraciones de hormonas esteroides androgénicas y estrogénicas, los endocrinólogos del comportamiento han estado particularmente interesados en la medida en que las diferencias de sexo conductual están mediadas por las hormonas. El proceso de llegar a ser femenino o masculino se llama diferenciación sexual. El primer paso en la diferenciación sexual ocurre en la fecundación. En los mamíferos, el óvulo (que siempre contiene un cromosoma X) puede ser fertilizado por un espermatozoide que porta un cromosoma Y o un cromosoma X; este proceso se llama determinación del sexo. El sexo cromosómico de los mamíferos homogaméticos (XX) es femenino; el sexo cromosómico de los mamíferos heterogaméticos (XY) es macho. El sexo cromosómico determina el sexo gonadal. Prácticamente toda la diferenciación sexual posterior es típicamente el resultado de la exposición diferencial a las hormonas esteroides gonadales. Así, el sexo gonadal determina el sexo hormonal, el cual regula el sexo morfológico. Las diferencias morfológicas en el sistema nervioso central, así como en algunos órganos efectores, como los músculos, conducen a diferencias de sexo conductual. El proceso de diferenciación sexual es complicado, y el potencial de errores está presente. La exposición perinatal a andrógenos es la causa más común de diferenciación sexual anómala entre las mujeres. La fuente de andrógeno puede ser interna (por ejemplo, secretada por las glándulas suprarrenales) o externa (por ejemplo, exposición a estrógenos ambientales). El síndrome de Turner resulta cuando el segundo cromosoma X falta o está dañado; estos individuos poseen ovarios disgénicos y no están expuestos a hormonas esteroides hasta la pubertad. Curiosamente, las mujeres con síndrome de Turner a menudo tienen memoria espacial deteriorada.
Las hembras se consideran el sexo “neutro”; se requieren pasos fisiológicos adicionales para la diferenciación masculina, y más pasos traen más posibilidades de errores en la diferenciación. Algunos ejemplos de diferenciación sexual anómala masculina incluyen la deficiencia de 5α-reductasa (en la que los individuos XY nacen con genitales ambiguos por falta de dihidrotestosterona y se crían como hembras, pero la masculinización ocurre durante la pubertad) y el síndrome de insensibilidad a andrógenos o TFM (en el que los individuos XY carecen de receptores para andrógenos y se desarrollan como hembras). Al estudiar a individuos que no caen pulcramente en las cajas dicóticas de mujeres o hombres y para quienes el proceso de diferenciación sexual es atípico, los endocrinólogos conductuales recogen pistas sobre el proceso de diferenciación sexual típica.
En última instancia, es posible que queramos saber cómo las hormonas median las diferencias de sexo en el cerebro y el comportamiento humanos (en la medida en que ocurren estas diferencias). Para comprender los mecanismos que subyacen a las diferencias de sexo en el cerebro y el comportamiento, volvemos al ejemplo del canto de los pájaros. Las aves proporcionan la mejor evidencia de que las diferencias de sexo conductual son el resultado de cambios estructurales inducidos hormonalmente en el cerebro (Goodson, Saldanha, Hahn, & Soma, 2005). A diferencia de los mamíferos, en los que las diferencias estructurales en los tejidos neurales no se han relacionado directamente con el comportamiento, las diferencias estructurales en los cerebros aviares se han relacionado directamente con un comportamiento sexual: el canto de los pájaros.
Varias regiones cerebrales en pájaros cantores muestran diferencias significativas de tamaño por sexo. Dos vías principales del circuito cerebral, (1) la vía motora de producción de canciones y (2) la vía de transmisión auditiva, han sido implicadas en el aprendizaje y producción del canto de los pájaros. Algunas partes de la ruta de producción de canciones de pinzones cebra machos son de 3 a 6 veces más grandes que las de los congéneres femeninos. El mayor tamaño de estas áreas cerebrales refleja que las neuronas en estos núcleos son más grandes, más numerosas y más separadas. Aunque la castración de aves macho adultas reduce el canto, no reduce el tamaño de los núcleos cerebrales controlando la producción de canto. De manera similar, el tratamiento con andrógenos de pinzones cebra hembras adultas no induce cambios ni en el canto ni en el tamaño de las regiones de control de la canción. Por lo tanto, los efectos activacionales de las hormonas esteroides no explican las diferencias de sexo en el comportamiento de canto o el tamaño del núcleo cerebral en los pinzones cebra. Las diferencias de sexo en estas estructuras son organizadas o programadas en el óvulo por estradiol (masculiniza) o la falta de esteroides (feminiza).
Tomados en conjunto, los estrógenos parecen ser necesarios para activar la maquinaria neuronal subyacente al sistema de canto en las aves. Los testículos de las aves producen principalmente andrógenos, que ingresan a la circulación. Los andrógenos ingresan a las neuronas que contienen aromatasa, lo que los convierte en estrógenos. En efecto, el cerebro es la principal fuente de estrógenos, que activan comportamientos masculinos en muchas especies de aves.
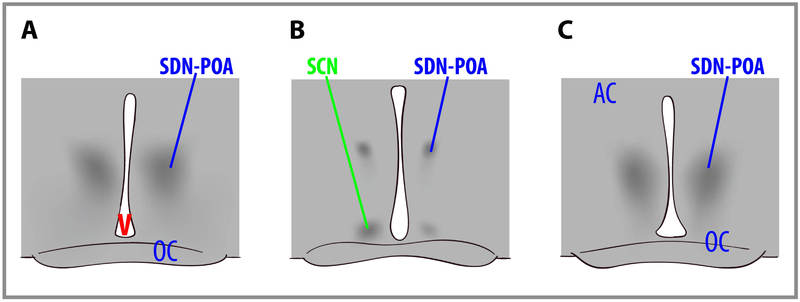
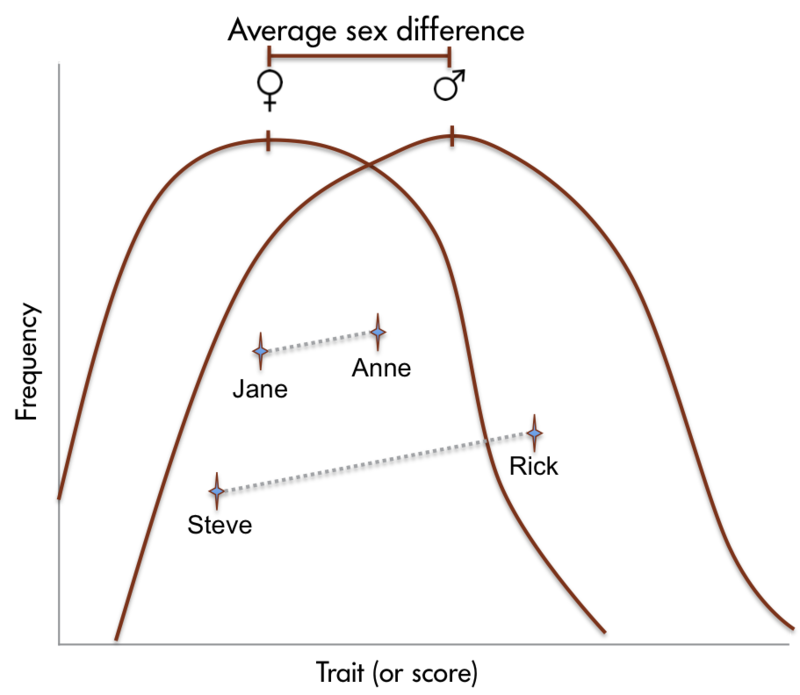
Las diferencias de sexo en el tamaño del cerebro humano se han reportado desde hace años. Más recientemente, se han descubierto diferencias de sexo en estructuras cerebrales específicas (Figura 1.5.2). También se han reportado diferencias de sexo en una serie de funciones cognitivas. Las hembras son generalmente más sensibles a la información auditiva, mientras que los machos son más sensibles a la información visual. Las hembras también suelen ser más sensibles que los machos al gusto y al aporte olfativo. Las mujeres muestran menos lateralización de las funciones cognitivas que los hombres. En promedio, las hembras generalmente sobresalen en habilidades verbales, perceptuales y motrices finas, mientras que los machos superan a las hembras en tareas cuantitativas y visuoespaciales, incluyendo lectura de mapas y búsqueda de dirección. Aunque se pueden documentar diferencias de sexo confiables, estas diferencias en la capacidad son leves. Es importante señalar que hay más variación dentro de cada sexo que entre los sexos para la mayoría de las habilidades cognitivas (Figura 1.5.3).
Comportamientos Agresivos
La posibilidad de comportamiento agresivo existe siempre que los intereses de dos o más individuos están en conflicto (Nelson, 2006). Es más probable que surjan conflictos por recursos limitados como territorios, alimentos y compañeros. Una interacción social decide qué animal obtiene acceso al recurso impugnado. En muchos casos, una postura sumisa o gesto por parte de un animal evita la necesidad de un combate real sobre un recurso. Los animales también pueden participar en exhibiciones de amenazas o combates ritualizados en los que se determine el dominio pero no se inflija ningún daño físico.
Existe evidencia circunstancial abrumadora de que las hormonas esteroides androgénicas median el comportamiento agresivo en muchas especies. Primero, coinciden las variaciones estacionales en las concentraciones plasmáticas de testosterona y las variaciones estacionales en la agresión. Por ejemplo, la incidencia de comportamiento agresivo alcanza su punto máximo para los ciervos machos en otoño, cuando están secretando altos niveles de testosterona. Segundo, las conductas agresivas aumentan en el momento de la pubertad, cuando los testículos se vuelven activos y aumentan las concentraciones sanguíneas de andrógenos. Los venados juveniles no participan en los combates durante la época de apareamiento. Tercero, en cualquier especie dada, los machos son generalmente más agresivos que las hembras. Esto es ciertamente cierto en el caso de los ciervos; en relación con los ciervos, las hembras rara vez muestran un comportamiento agresivo, y sus raros actos agresivos son cualitativamente diferentes del comportamiento agresivo de los machos agresivos. Finalmente, la castración generalmente reduce la agresión en los hombres, y la terapia de reemplazo de testosterona restaura la agresión a los niveles previos a la castración. Existen algunas excepciones interesantes a estas observaciones generales que están fuera del alcance de este módulo.
Como se mencionó, los machos son generalmente más agresivos que las hembras. Ciertamente, los machos humanos son mucho más agresivos que las hembras. Muchos más hombres que mujeres son condenados por delitos violentos en Norteamérica. Las diferencias de sexo en la agresividad humana aparecen muy tempranas. A todas las edades a lo largo de los años escolares, muchos más niños que niñas inician agresiones físicas. Casi todos reconocerán la existencia de esta diferencia de sexo, pero asignar una causa a las diferencias de sexo conductual en humanos siempre provoca mucho debate. Es posible que los niños sean más agresivos que las niñas porque los andrógenos promueven un comportamiento agresivo y los niños tienen mayores concentraciones de andrógenos en sangre que las niñas. Es posible que los niños y niñas difieran en su agresividad porque los cerebros de los niños están expuestos a los andrógenos prenatalmente y el “cableado” de sus cerebros se organiza así de una manera que facilita la expresión de la agresión. También es posible que los niños sean alentados y las niñas sean desalentadas por la familia, compañeros u otras personas de actuar de manera agresiva. Estas tres hipótesis no son mutuamente excluyentes, pero es extremadamente difícil discriminar entre ellas para dar cuenta de las diferencias de sexo en la agresividad humana.

¿Qué tipo de estudios serían necesarios para evaluar estas hipótesis? Por lo general, es difícil separar las influencias del ambiente y la fisiología en el desarrollo del comportamiento en los humanos. Por ejemplo, niños y niñas difieren en su juego rudo a una edad muy temprana, lo que sugiere una influencia fisiológica temprana en la agresión. Sin embargo, los padres interactúan con su descendencia masculina y femenina de manera diferente; suelen jugar más bruscamente con bebés varones que con hembras, lo que sugiere que la diferencia de sexo en la agresividad se aprende parcialmente. Esta diferencia en el estilo de interacción parental es evidente para la primera semana de vida. Debido a estas complejidades en los factores que influyen en el comportamiento humano, se ha perseguido el estudio de los efectos hormonales sobre el comportamiento diferenciado por sexo en animales no humanos, para lo cual las influencias ambientales pueden mantenerse relativamente constantes. A menudo se utilizan modelos animales para los que la diferenciación sexual ocurre posnatalmente para que este proceso pueda ser fácilmente manipulado experimentalmente.
Nuevamente, con el modelo animal apropiado, podemos abordar las preguntas planteadas anteriormente: ¿La diferencia de sexo en la agresión se debe a mayores concentraciones de andrógenos en la sangre adulta en los machos que en las hembras, o son los machos más agresivos que las hembras porque sus cerebros están organizados de manera diferente por las hormonas perinatales? ¿Los machos suelen ser más agresivos que las hembras debido a una interacción de las concentraciones tempranas y actuales de andrógenos en sangre? Si los ratones machos son castrados antes de su sexto día de vida, luego tratados con propionato de testosterona en la edad adulta, muestran bajos niveles de agresión. De manera similar, las hembras ovariectomizadas antes de su sexto día pero administradas andrógenos en la edad adulta no expresan niveles de agresión similares a los del hombre. El tratamiento de machos o hembras perinatalmente gonadectomizados con testosterona antes de su sexto día de vida y también en la edad adulta da como resultado un nivel de agresión similar al observado en ratones machos típicos. Así, en ratones, la proclividad para que los machos actúen de manera más agresiva que las hembras está organizada perinatalmente por los andrógenos pero también requiere la presencia de andrógenos después de la pubertad para que se expresen plenamente. En otras palabras, la agresión en ratones machos está organizada y activada por los andrógenos. La exposición a la testosterona en la edad adulta sin organización previa del cerebro por las hormonas esteroides no evoca niveles típicos de agresión masculina. El control hormonal del comportamiento agresivo en ratones domésticos es, por lo tanto, similar a la mediación hormonal del comportamiento de apareamiento masculino heterosexual en otras especies de roedores. El comportamiento agresivo es organizado y activado por andrógenos en muchas especies, incluyendo ratas, hámsters, ratones de campo, perros y posiblemente algunas especies de primates.
Comportamientos de los padres
El comportamiento de los padres puede considerarse como cualquier comportamiento que contribuya directamente a la supervivencia de los óvulos fecundados o crías que han abandonado el cuerpo de la hembra. Hay muchos patrones de cuidado parental de mamíferos. El estado de desarrollo del recién nacido es un factor importante que impulsa el tipo y la calidad del cuidado parental en una especie. La atención materna es mucho más común que la atención paterna. La gran mayoría de las investigaciones sobre los correlatos hormonales del comportamiento parental de los mamíferos se han realizado en ratas. Las ratas tienen crías altriciales y las madres realizan un grupo de comportamientos maternos estereotipados, que incluyen la construcción de nidos, agacharse sobre los cachorros para permitir la lactancia y proporcionar calidez, recuperación de crías y aumento de la agresión dirigida a los intrusos. Si expone ratas hembra (o machos) no preñadas a crías, su reacción más común es acurrucarse lejos de ellas. Las ratas evitan cosas nuevas (neofobia). No obstante, si expones ratas adultas a crías todos los días, pronto empiezan a comportarse de manera materna. Este proceso se llama concaveación o sensibilización y parece servir para reducir el miedo de las ratas adultas a los cachorros.
Por supuesto, una nueva madre necesita actuar como materna tan pronto como llegue su descendencia, no en una semana. El inicio del comportamiento materno en ratas está mediado por hormonas. Varios métodos de estudio, como la eliminación hormonal y la terapia de reemplazo, se han utilizado para determinar los correlatos hormonales del comportamiento materno de la rata. Una rápida disminución de las concentraciones sanguíneas de progesterona al final del embarazo después de altas concentraciones sostenidas de esta hormona, en combinación con altas concentraciones de estradiol y probablemente prolactina y oxitocina, induce a las ratas hembra a comportarse materno casi de inmediato en presencia de crías. Este patrón de hormonas en el parto anula la respuesta de miedo habitual de las ratas adultas hacia las crías, y permite el inicio de la conducta materna. Así, el llamado “instinto” materno requiere hormonas para aumentar la tendencia de acercamiento y disminuir la tendencia a la evitación. Las cepas de laboratorio de ratones y ratas suelen ser dóciles, pero las madres pueden ser bastante agresivas con los animales que se aventuran demasiado cerca de su camada. La progesterona parece ser la principal hormona que induce esta agresión materna en roedores, pero existen diferencias de especies. No se ha descrito ni probado adecuadamente el papel de la agresión materna en el comportamiento de las mujeres.

Una serie de elegantes experimentos de Alison Fleming y sus colaboradores estudiaron los correlatos endocrinos del comportamiento de las madres humanas así como los correlatos endocrinos de las actitudes maternas expresadas en cuestionarios de autoinforme. Las respuestas como dar palmaditas, abrazar o besar al bebé se llamaron comportamientos afectuosos; hablar, cantar o arrullar al bebé se consideraron comportamientos vocales. Tanto los comportamientos afectuosos como los vocales se consideraron conductas de abordaje. También se registraron actividades básicas de cuidado, como cambiar pañales y eructar a los infantes. En estos estudios no se encontró relación entre las concentraciones hormonales y la capacidad de respuesta materna, medida por cuestionarios de actitud. Por ejemplo, la mayoría de las mujeres mostraron una autoimagen positiva creciente durante el embarazo temprano que se sumergió durante la segunda mitad del embarazo, pero se recuperó después del parto. Una caída relacionada en los sentimientos de compromiso materno ocurrió durante el embarazo tardío, pero rebotó sustancialmente después del nacimiento en la mayoría de las mujeres. Sin embargo, cuando se comparó el comportamiento, más que las respuestas al cuestionario, con las concentraciones hormonales, surgió una historia diferente. Las concentraciones plasmáticas de cortisol se asociaron positivamente con comportamientos de abordaje. Es decir, las mujeres que presentaban altas concentraciones de cortisol en sangre, en muestras obtenidas inmediatamente antes o después de amamantar, realizaban comportamientos más afectuosos físicamente y platicaban con más frecuencia con sus bebés que las madres con bajas concentraciones de cortisol. Análisis adicionales de este estudio revelaron que la correlación fue aún mayor para las madres que habían reportado respeto materno positivo (sentimientos y actitudes) durante la gestación. De hecho, casi la mitad de la variación en el comportamiento materno entre las mujeres podría explicarse por las concentraciones de cortisol y las actitudes maternas positivas durante el embarazo.
Presumiblemente, el cortisol no induce comportamientos maternos directamente, sino que puede actuar indirectamente sobre la calidad de la atención materna al evocar un aumento en el nivel general de excitación de la madre, aumentando así su capacidad de respuesta a las señales generadas por el bebé. Las nuevas madres con altas concentraciones de cortisol también se sintieron más atraídas por los olores de sus bebés, fueron superiores en la identificación de sus bebés y generalmente encontraron señales de bebés muy atractivas (Fleming, Steiner, & Corter, 1997).
El área preóptica medial es crítica para la expresión del comportamiento materno de la rata. La amígdala parece inhibir tónicamente la expresión del comportamiento materno. Las ratas adultas temen a los cachorros, una respuesta que aparentemente está mediada por información quimiosensorial. Las lesiones de la amígdala o vías sensoriales aferentes desde el órgano vomeronasal hasta la amígdala desinhiben la expresión del comportamiento materno. Las hormonas o sensibilización probablemente actúan para desinhibir la amígdala, permitiendo así la ocurrencia de la conducta materna. Aunque se han establecido correlaciones, la evidencia directa de cambios estructurales cerebrales en madres humanas sigue sin especificar (Fleming & Gonzalez, 2009).
Considerados en conjunto, hay muchos ejemplos de hormonas que influyen en el comportamiento y de comportamiento que retroalimentan para influir en la secreción hormonal. Cada vez se descubren más ejemplos de interacciones hormona-comportamiento, incluyendo hormonas en la mediación de la ingesta de alimentos y líquidos, interacciones sociales, equilibrio de sal, aprendizaje y memoria, afrontamiento del estrés, así como psicopatología que incluye depresión, trastornos de ansiedad, trastornos alimentarios, depresión posparto, y depresión estacional. Investigaciones adicionales deberían revelar cómo están mediadas estas interacciones hormona-comportamiento.
Recursos Externos
- Libro: Adkins-Regan, E. (2005). Hormonas y comportamiento social animal. Princeton, NJ: Prensa de la Universidad de Princeton.
- Libro: Playa, F. A. (1948). Hormonas y comportamiento. Nueva York: Paul Hoeber.
- Libro: Playa, F. A. (1975). Endocrinología conductual: Una disciplina emergente. Científico Americano, 63:178—187.
- Libro: Nelson, R. J. (2011). Una introducción a la endocrinología conductual (4ª ed.). Sunderland, MA: Asociados Sinauer.
- Libro: Pfaff, D. W. (2009). Hormonas, cerebro y comportamiento (2ª ed.). Nueva York: Prensa Académica.
- Libro: Pfaff, D. W., Phillips, I. M., & Rubin, R. T. (2005). Principios de las relaciones hormona/conducta. Nueva York: Prensa Académica.
- Video: Endocrinología Video (Playlist) - Esta lista de reproducción de YouTube contiene muchos videos útiles sobre la biología de las hormonas, incluida la reproducción y el comportamiento. Este sería un recurso útil para los estudiantes que luchan con la síntesis hormonal, la reproducción, la regulación de las funciones biológicas y las vías de señalización.
- https://www.youtube.com/playlist?list=PLqTetbgey0aemiTfD8QkMsSUq8hQzv-vA
- Video: Paul Zak: Confianza, moralidad -y oxitocina- Esta plática de Ted explora los papeles de la oxitocina en el cuerpo. Paul Zak analiza las funciones biológicas de la oxitocina, como la lactancia, así como las posibles funciones conductuales, como la empatía.
- Video: Diferenciación Sexual- Este video analiza la diferenciación gonadal, incluyendo el papel de los andrógenos en el desarrollo de rasgos masculinos.
- Video: Explicación del cerebro adolescente- Este es un gran video que explica los roles de las hormonas durante la pubertad.
- Web: Sociedad de Neuroendocrinología Conductual - Este sitio web contiene recursos sobre noticias de actualidad e investigación en el campo de la neuroendocrinología.
- http://sbn.org/home.aspx
Preguntas de Discusión
- ¿Cuáles son algunos de los problemas asociados al intento de determinar la causalidad en una interacción horno—comportamiento? ¿Cuáles son las mejores formas de abordar estos problemas?
- Las hormonas provocan cambios en las tasas de procesos celulares o en la morfología celular. ¿Cuáles son algunas formas en que estos cambios celulares inducidos por hormonas podrían producir teóricamente cambios profundos en el comportamiento?
- Enumere y describa algunas diferencias de sexo conductual que haya notado entre niños y niñas. ¿Qué causa que niñas y niños elijan diferentes juguetes? ¿Crees que las diferencias de sexo que has notado surgen de causas biológicas o se aprenden? ¿Cómo harías para establecer tus opiniones como un hecho?
- ¿Por qué es inapropiado referirse a los andrógenos como hormonas “masculinas” y a los estrógenos como hormonas “femeninas”?
- Imagínese que descubrió que los cerebros de los arquitectos eran diferentes a los de los no arquitectos, específicamente, que los “núcleos drawstraightem” del lóbulo temporal derecho se ampliaron en arquitectos en comparación con los no arquitectos. ¿Argumentaría que los arquitectos estaban destinados a ser arquitectos por su organización cerebral o que la experiencia como arquitecto les cambió el cerebro? ¿Cómo resolverías este problema?
El vocabulario
- 5α-reductasa
- Una enzima requerida para convertir la testosterona en 5α-dihidrotestosterona.
- Agresión
- Una forma de interacción social que incluye amenazas, ataques y combates.
- Aromatasa
- Enzima que convierte los andrógenos en estrógenos.
- Sexo cromosómico
- El sexo de un individuo determinado por los cromosomas sexuales (típicamente XX o XY) recibidos al momento de la fertilización.
- Defeminización
- La eliminación del potencial de rasgos femeninos.
- Desmasculinización
- La eliminación del potencial de rasgos masculinos.
- Dihidrotestosterona (DHT)
- Un andrógeno primario que es un producto esteroide androgénico de la testosterona y se une fuertemente a los receptores de andrógenos.
- Glándula endocrina
- Glándula sin conductos de la cual se liberan hormonas al sistema sanguíneo en respuesta a señales biológicas específicas.
- Estrógeno
- Cualquiera de la clase C18 de hormonas esteroides, llamado así por las propiedades generadoras de estrus en las hembras. Los estrógenos biológicamente importantes incluyen estradiol y estriol.
- Feminización
- La inducción de rasgos femeninos.
- Sexo gonadal
- El sexo de un individuo determinado por la posesión de ovarios o testículos. Las hembras tienen ovarios, mientras que los machos tienen testículos.
- Hormona
- Un mensajero químico orgánico liberado de las células endocrinas que viaja a través de la sangre para interactuar con las células diana a cierta distancia para provocar una respuesta biológica.
- Masculinización
- La inducción de rasgos masculinos.
- Comportamiento materno
- Comportamiento parental realizado por la madre u otra hembra.
- Neurotransmisor
- Un mensajero químico que viaja entre neuronas para proporcionar comunicación. Algunos neurotransmisores, como la norepinefrina, pueden filtrarse al sistema sanguíneo y actuar como hormonas.
- Oxitocina
- Una hormona peptídica secretada por la glándula pituitaria para desencadenar la lactancia, así como el vínculo social.
- Comportamiento de los padres
- Comportamientos realizados en relación con la propia descendencia que contribuyen directamente a la supervivencia de esas crías
- Comportamiento paterno
- Comportamiento parental realizado por el padre u otro varón.
- Progesterona
- Progestina primaria que interviene en conductas de embarazo y apareamiento.
- Progestina
- Una clase de hormonas esteroides C21 nombradas por sus efectos progestacionales (de apoyo al embarazo). La progesterona es una progestina común.
- Prohormona
- Una molécula que puede actuar como una hormona en sí misma o convertirse en otra hormona con diferentes propiedades. Por ejemplo, la testosterona puede servir como hormona o como prohormona para la dihidrotestosterona o el estradiol.
- Prolactina
- Una hormona proteica que está altamente conservada en todo el reino animal. Tiene muchas funciones biológicas asociadas con la reproducción y acciones sinérgicas con las hormonas esteroides.
- Receptor
- Una estructura química en la superficie celular o dentro de una célula que tiene afinidad por una configuración química específica de una hormona, neurotransmisor u otro compuesto.
- Determinación del sexo
- El punto en el que un individuo comienza a desarrollarse ya sea como varón o hembra. En animales que tienen cromosomas sexuales, esto ocurre en la fertilización. Las hembras son XX y los machos son XY. Todos los óvulos llevan cromosomas X, mientras que los espermatozoides pueden portar cromosomas X o Y. Así, son los machos los que determinan el sexo de la descendencia.
- Diferenciación sexual
- Proceso por el cual los individuos desarrollan las características asociadas al ser masculino o femenino. La exposición diferencial a los esteroides gonadales durante el desarrollo temprano provoca la diferenciación sexual de varias estructuras, incluido el cerebro.
- Célula objetivo
- Una célula que tiene receptores para un mensajero químico específico (hormona o neurotransmisor).
- Testosterona
- El andrógeno primario secretado por los testículos de la mayoría de los animales vertebrados, incluidos los hombres.
Referencias
- Alexander, G. M. & Hines, M. (2002). Diferencias de sexo en respuesta a juguetes infantiles en primates no humanos (Cercopithecus aethiops sabaeus). Evolución y Comportamiento Humano, 23, 467—479.
- Berenbaum, S. A., Martin, C. L., Hanish, L. D., Briggs, P. T., & Fabes, R. A. (2008). Diferencias sexuales en el juego infantil. En J. B. Becker, K. J. Berkley, N. Geary, E. Hampson, J. Herman, & E. Young (Eds.), Diferencias sexuales en el cerebro: De los genes al comportamiento. Nueva York: Oxford University Press.
- Dabbs, J. M. (2000). Héroes, pícaros y amantes: Testosterona y comportamiento. Columbus, OH: McGraw Hill.
- Fleming, A. S., & González, A. (2009). Neurobiología de la atención materna humana. En P. T. Ellison & P. B. Gray (Eds.), Endocrinología de las relaciones sociales (pp. 294—318). Cambridge, MA: Prensa de la Universidad de Harvard.
- Fleming, A. S., Steiner, M., & Corter, C. (1997). Cortisol, hedónica y capacidad de respuesta materna en madres humanas. Hormonas y Conducta, 32, 85—98.
- Goodson, J. L., Saldanha, C. J., Hahn, T. P., Soma, K. K. (2005). Avances recientes en neuroendocrinología conductual: Insights from studies on birds. Hormonas y Conducta, 48, 461—73.
- Kidd, K. A., Blanchfield, P. J., Mills, K. H., Palace, V. P., Evans, R. E. Lazorchak, J. M. & Flick, R. (2007). Colapso de una población de peces tras la exposición a un estrógeno sintético. Actas de la Academia Nacional de Ciencias,104, 8897—8901.
- Nelson, R. J. (Ed.) (2006). Biología de la agresión. Nueva York: Oxford University Press.
- Nelson, R.J. (2011). Una introducción a la endocrinología conductual (4ª ed.). Sunderland, MA: Asociados Sinauer.
- Phoenix, C. H., Goy, R. W., Gerall, A. A., & Young, W. C. (1959). Acción organizadora del propionato de testosterona administrado prenatalmente sobre los tejidos mediando el comportamiento de apareamiento en el conejillo de indias hembra. Endocrinología, 65:369—382.
- van Anders, S., Hamilton, L., Schmidt, N., & Watson, N. (2007). Asociaciones entre secreción de testosterona y actividad sexual en mujeres. Hormonas y Comportamiento, 51, 477—82.