
4.1: Neurociencia Afectiva
- Page ID
- 144023

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Universidad de Nueva Gales del Sur
Este módulo proporciona una breve descripción de la neurociencia de la emoción. Integra hallazgos de investigaciones humanas y animales para describir las redes cerebrales y los neurotransmisores asociados involucrados en los sistemas afectivos básicos.
objetivos de aprendizaje
- Definir la neurociencia afectiva.
- Describir las técnicas de neurociencia utilizadas para estudiar las emociones en humanos y animales.
- Nombrar cinco sistemas emocionales y sus estructuras neuronales y neurotransmisores asociados.
- Dar ejemplos de sustancias químicas exógenas (por ejemplo, drogas) que influyen en los sistemas afectivos, y discutir sus efectos.
- Discutir múltiples funciones afectivas de la amígdala y el núcleo accumbens.
- Nombrar varias emociones humanas específicas y discutir su relación con los sistemas afectivos de animales no humanos.
Neurociencia Afectiva: ¿Qué es?
La neurociencia afectiva examina cómo el cerebro crea respuestas emocionales. Las emociones son fenómenos psicológicos que involucran cambios en el cuerpo (p. ej., expresión facial), cambios en la actividad del sistema nervioso autónomo, estados sensoriales (respuestas subjetivas) e instos a actuar de maneras específicas (motivaciones; Izard, 2010). La neurociencia afectiva tiene como objetivo comprender cómo la materia (estructuras cerebrales y sustancias químicas) crea uno de los aspectos más fascinantes de la mente, las emociones. La neurociencia afectiva utiliza medidas imparciales y observables que proporcionan evidencia creíble a otras ciencias y laicos sobre la importancia de las emociones. También conduce a tratamientos de base biológica para trastornos afectivos (p. ej., depresión).

El cerebro humano y sus respuestas, incluyendo las emociones, son complejos y flexibles. En comparación, los animales no humanos poseen sistemas nerviosos más simples y respuestas emocionales más básicas. Las técnicas invasivas de neurociencia, como la implantación de electrodos, la lesión y la administración de hormonas, se pueden usar más fácilmente en animales que en humanos. La neurociencia humana debe basarse principalmente en técnicas no invasivas como la electroencefalografía (EEG) y la resonancia magnética funcional (fMRI), y en estudios de individuos con lesiones cerebrales causadas por accidente o enfermedad. Así, la investigación animal proporciona modelos útiles para entender los procesos afectivos en humanos. Los circuitos afectivos encontrados en otras especies, particularmente mamíferos sociales como ratas, perros y monos, funcionan de manera similar a las redes afectivas humanas, aunque los cerebros de los animales no humanos son más básicos.
En los humanos, las emociones y sus sistemas neuronales asociados tienen capas adicionales de complejidad y flexibilidad. En comparación con los animales, los humanos experimentan una gran variedad de emociones matizadas y a veces contradictorias. Los humanos también responden a estas emociones de formas complejas, de tal manera que las metas conscientes, los valores y otras cogniciones influyen en el comportamiento además de las respuestas emocionales. Sin embargo, en este módulo nos centramos en las similitudes entre organismos, más que en las diferencias. A menudo usamos el término “organismo” para referirnos al individuo que está experimentando una emoción o mostrando evidencia de activaciones neuronales particulares. Un organismo podría ser una rata, un mono o un ser humano.
En todas las especies, las respuestas emocionales se organizan en torno a la supervivencia y las necesidades reproductivas del organismo. Las emociones influyen en la percepción, la cognición y el comportamiento para ayudar a los organismos a sobrevivir y prosperar (Farb, Chapman y Anderson, 2013). Las redes de estructuras en el cerebro responden a diferentes necesidades, con cierta superposición entre diferentes emociones. Las emociones específicas no se localizan en una sola estructura del cerebro. En cambio, las respuestas emocionales implican redes de activación, con muchas partes del cerebro activadas durante cualquier proceso emocional. De hecho, los circuitos cerebrales involucrados en las reacciones emocionales incluyen casi todo el cerebro (Berridge & Kringelbach, 2013). Los circuitos cerebrales ubicados en lo profundo del cerebro por debajo de la corteza cerebral son los principales responsables de generar emociones básicas (Berridge & Kringelbach, 2013; Panksepp & Biven, 2012). En el pasado, la atención de la investigación se centró en estructuras cerebrales específicas que se revisarán aquí, pero investigaciones futuras pueden encontrar que áreas adicionales del cerebro también son importantes en estos procesos.
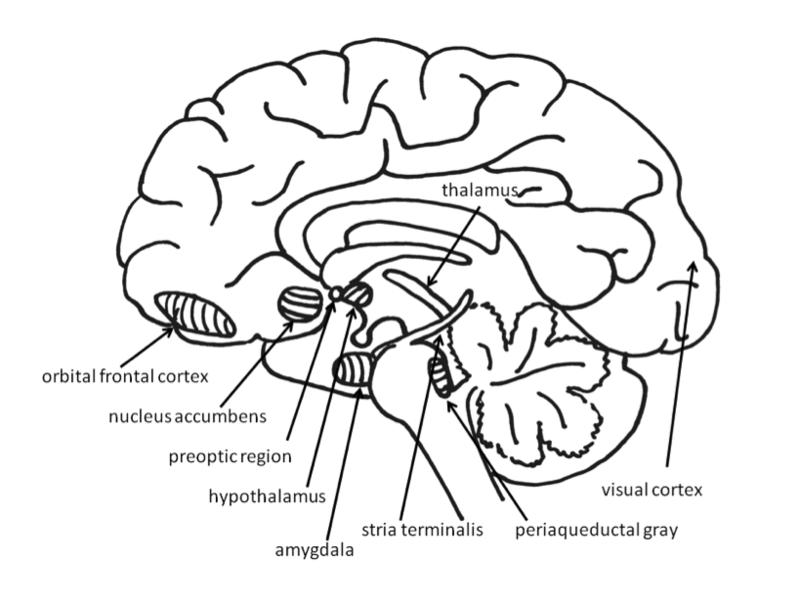
Emociones Básicas
Deseo: Los sistemas neuronales de la búsqueda de recompensas
Uno de los sistemas neuronales afectivos más importantes se relaciona con los sentimientos de deseo, o el apetito por las recompensas. Los investigadores se refieren a estos procesos apetitivos usando términos como “querer” (Berridge & Kringelbach, 2008), “seeking” (Panksepp & Biven, 2012) o “sensibilidad de activación conductual” (Gray, 1987). Cuando se despierta el sistema apetitivo, el organismo muestra entusiasmo, interés y curiosidad. Estos circuitos neuronales motivan al animal a moverse por su entorno en busca de recompensas como alimentos apetitosos, parejas sexuales atractivas y otros estímulos placenteros. Cuando el sistema apetitivo está subexcitado, el organismo aparece deprimido e indefenso.
Mucha evidencia de las estructuras involucradas en este sistema proviene de la investigación animal usando estimulación cerebral directa. Cuando se implanta un electrodo en el hipotálamo lateral o en regiones corticales o mesencefálicas a las que está conectado el hipotálamo, los animales presionarán una palanca para entregar estimulación eléctrica, sugiriendo que encuentran placentera la estimulación. Las regiones en el sistema de deseo también incluyen la amígdala, el núcleo accumbens y la corteza frontal (Panksepp & Biven, 2012). El neurotransmisor dopamina, producido en los circuitos mesolímbico y mesocortical de dopamina, activa estas regiones. Crea una sensación de emoción, sentido y anticipación. Estas estructuras también son sensibles a drogas como la cocaína y las anfetaminas, sustancias químicas que tienen efectos similares a la dopamina (Panksepp & Biven, 2012).

La investigación tanto en humanos como en animales no humanos muestra que la corteza frontal izquierda (en comparación con la corteza frontal derecha) es más activa durante las emociones apetitivas como el deseo y el interés. Los investigadores señalaron por primera vez que las personas que habían sufrido daños en la corteza frontal izquierda desarrollaron depresión, mientras que aquellas con daño en la corteza frontal derecha desarrollaron manía (Goldstein, 1939). La relación entre la activación frontal izquierda y las emociones relacionadas con el enfoque se ha confirmado en individuos sanos usando EEG y fMRI (Berkman & Lieberman, 2010). Por ejemplo, el aumento de la activación frontal izquierda ocurre en bebés de 2 a 3 días de edad cuando se les coloca sacarosa en la lengua (Fox & Davidson, 1986), y en adultos hambrientos al ver imágenes de postres deseables (Gable & Harmon-Jones, 2008). Además, se ha encontrado que una mayor actividad frontal izquierda en situaciones apetitivas se relaciona con la dopamina (Wacker, Mueller, Pizzagalli, Hennig, & Stemmler, 2013).
“Gusto”: Los circuitos neuronales del placer y el disfrute
Sorprendentemente, la cantidad de deseo que siente un individuo hacia una recompensa no necesita corresponder a cuánto le gusta esa recompensa. Esto se debe a que las estructuras neuronales involucradas en el disfrute de las recompensas son diferentes de las estructuras involucradas en el deseo de las recompensas. El “gusto” (por ejemplo, el disfrute de un líquido dulce) se puede medir en bebés y animales no humanos midiendo la velocidad de lamer, protuberancias en la lengua y expresiones faciales felices, mientras que “querer” (deseo) se muestra por la voluntad de trabajar duro para obtener una recompensa (Berridge y Kringelbach, 2008). El gusto se ha distinguido de querer en la investigación sobre temas como el abuso de drogas. Por ejemplo, los drogadictos a menudo desean drogas incluso cuando saben que las disponibles no proporcionarán placer (Stewart, de Wit, & Eikelboom, 1984).
La investigación sobre el gusto se ha centrado en una pequeña área dentro del núcleo accumbens y en la mitad posterior del palidum ventral. Estas regiones cerebrales son sensibles a los opioides y endocannabinoides. La estimulación de otras regiones del sistema de recompensa aumenta el querer, pero no aumenta el gusto, y en algunos casos incluso disminuye el gusto. La investigación sobre la distinción entre deseo y disfrute contribuye a la comprensión de la adicción humana, particularmente por qué los individuos a menudo continúan buscando frenéticamente recompensas como la cocaína, los opiáceos, el juego o el sexo, incluso cuando ya no experimentan placer al obtener estas recompensas debido a habituación.
La experiencia del placer también involucra la corteza orbitofrontal. Las neuronas en esta región se incendian cuando los monos prueban, o simplemente ven imágenes de, alimentos deseables. En los humanos, esta región es activada por estímulos agradables que incluyen dinero, olores agradables y caras atractivas (Gottfried, O'Doherty & Dolan, 2002; O'Doherty, Deichmann, Critchley, & Dolan, 2002; O'Doherty, Kringelbach, Rolls, Hornak, & Andrews, 2001; O'Doherty, Winston, Critchley, Perrett, Burt, & Andrews, 2001; O'Doherty, Winston, Critchley, Perrett, Burt, & ; Dolan, 2003).
Miedo: El sistema neural de la congelación y la huida

El miedo es una emoción desagradable que motiva a evitar situaciones potencialmente dañinas. La ligera estimulación de las áreas relacionadas con el miedo en el cerebro hace que los animales se congelen, mientras que la estimulación intensa hace que huyan. El circuito del miedo se extiende desde la amígdala central hasta el gris periacueductal en el mesencéfalo. Estas estructuras son sensibles al glutamato, factor liberador de corticotrofina, hormona adreno-cortico-trófica, colecistoquinina y varios neuropéptidos diferentes. Las benzodiazepinas y otros tranquilizantes inhiben la activación en estas áreas (Panksepp & Biven, 2012).
El papel de la amígdala en las respuestas al miedo ha sido ampliamente estudiado. Quizás porque el miedo es tan importante para la supervivencia, dos vías envían señales a la amígdala desde los órganos sensoriales. Cuando un individuo ve una serpiente, por ejemplo, la información sensorial viaja del ojo al tálamo y luego a la corteza visual. La corteza visual envía la información sobre a la amígdala, provocando una respuesta de miedo. Sin embargo, el tálamo también envía rápidamente la información directamente a la amígdala, para que el organismo pueda reaccionar antes de percibir conscientemente a la serpiente (LeDoux, Farb, & Ruggiero, 1990). La vía desde el tálamo hasta la amígdala es rápida pero menos precisa que la ruta más lenta desde la corteza visual. El daño a la amígdala o áreas del hipocampo ventral interfiere con el condicionamiento del miedo tanto en humanos como en animales no humanos (LeDoux, 1996).
Rabia: Los circuitos de la ira y el ataque
La ira o la rabia es una emoción excitante y desagradable que motiva a los organismos a acercarse y atacar (Harmon-Jones, Harmon-Jones, & Price, 2013). La ira se puede evocar a través de la frustración de la meta, el dolor físico o la restricción física. En los animales territoriales, la ira es provocada por un extraño que ingresa al territorio natal del organismo (Blanchard & Blanchard, 2003). Las redes neuronales para la ira y el miedo están cerca unas de otras, pero separadas (Panksepp & Biven, 2012). Se extienden desde la amígdala medial, a través de partes específicas del hipotálamo, hasta el gris periacueductal del mesencéfalo. Los circuitos de ira están vinculados a los circuitos apetitivos, de tal manera que la falta de una recompensa anticipada puede provocar rabia. Además, cuando los humanos se enojan, muestran una mayor activación cortical frontal izquierda, apoyando la idea de que la ira es una emoción relacionada con el enfoque (Harmon-Jones et al., 2013). Los neurotransmisores involucrados en la rabia aún no se conocen bien, pero la Sustancia P puede jugar un papel importante (Panksepp & Biven, 2012). Otros neuroquímicos que pueden estar involucrados en la ira incluyen la testosterona (Peterson & Harmon-Jones, 2012) y arginina-vasopresina (Heinrichs, von Dawans, & Domes, 2009). Varios químicos inhiben el sistema de rabia, incluyendo opioides y altas dosis de antipsicóticos, como la clorpromazina (Panksepp & Biven, 2012).
Amor: Los sistemas neuronales de cuidado y apego

Para los animales sociales como los humanos, el apego a otros miembros de la misma especie produce las emociones positivas del apego: el amor, los sentimientos cálidos y el afecto. Las emociones que motivan el comportamiento nutritivo (por ejemplo, el cuidado materno) son distinguibles de aquellas que motivan mantenerse cerca de una figura de apego para recibir atención y protección (por ejemplo, apego infantil). Las regiones importantes para la crianza materna incluyen el área preóptica dorsal (Numan & Insel, 2003) y el núcleo de lecho de la estría terminal (Panksepp, 1998). Estas regiones se superponen con las áreas involucradas en el deseo sexual, y son sensibles a algunos de los mismos neurotransmisores, incluyendo oxitocina, arginina-vasopresina y opioides endógenos (endorfinas y encefalinas).
Duelo: Las redes neuronales de la soledad y el pánico
Las redes neuronales involucradas en el apego infantil también son sensibles a la separación. Estas regiones producen las emociones dolorosas del dolor, el pánico y la soledad. Cuando los humanos infantiles u otros mamíferos infantiles se separan de sus madres, producen vocalizaciones de angustia o llanto. Los circuitos de unión son aquellos que hacen que los organismos produzcan vocalizaciones de socorro cuando se estimulan eléctricamente.
El sistema de fijación comienza en el gris periacueductal mesencéfalo, muy cerca de la zona que produce respuestas físicas de dolor, lo que sugiere que puede haberse originado en los circuitos del dolor (Panksepp, 1998). La angustia por separación también se puede evocar estimulando el tálamo dorsomedial, el tabique ventral, la región preóptica dorsal y las áreas en el núcleo de la cama de la estría terminal (cerca de los circuitos sexuales y maternos; Panksepp, Normansell, Herman, Bishop, & Crepeau, 1988).
Estas regiones son sensibles a opiáceos endógenos, oxitocina y prolactina. Todos estos neurotransmisores previenen la angustia por separación. Los opiáceos como la morfina y la heroína, así como la nicotina, producen artificialmente sentimientos de placer y gratificación, similares a los que normalmente se producen durante las interacciones sociales positivas. Esto puede explicar por qué estas drogas son adictivas. Los ataques de pánico parecen ser una forma intensa de angustia por separación desencadenada por el sistema de apego, y el pánico puede ser efectivamente aliviado por los opiáceos. La testosterona también reduce la angustia por separación, tal vez al reducir las necesidades de apego. Consistente con esto, los ataques de pánico son más comunes en mujeres que en hombres.
Plasticidad: las experiencias pueden alterar el cerebro

Las respuestas de regiones neuronales específicas pueden ser modificadas por la experiencia. Por ejemplo, la cáscara frontal del núcleo accumbens generalmente está involucrada en comportamientos apetitivos, como comer, y la concha trasera generalmente está involucrada en comportamientos defensivos temerosos (Reynolds & Berridge, 2001, 2002). La investigación que utiliza neuroimagen humana también ha revelado esta distinción front—back en las funciones del núcleo accumbens (Seymour, Daw, Dayan, Singer, & Dolan, 2007). Sin embargo, cuando las ratas están expuestas a ambientes estresantes, sus regiones generadoras de miedo se expanden hacia el frente, llenando casi 90% de la cáscara del núcleo accumbens. Por otro lado, cuando las ratas están expuestas a ambientes domésticos preferidos, sus regiones generadoras de miedo se contraen y las regiones apetitivas se expanden hacia el dorso, llenando aproximadamente el 90% de la concha (Reynolds & Berridge, 2008).
Las estructuras cerebrales tienen múltiples funciones
Si bien gran parte de la investigación en neurociencia afectiva ha enfatizado estructuras enteras, como la amígdala y el núcleo accumbens, es importante señalar que muchas de estas estructuras son referidas con mayor precisión como complejos. Incluyen distintos grupos de núcleos que realizan diferentes tareas. En la actualidad, las técnicas de neuroimagen humana como la fMRI son incapaces de examinar la actividad de núcleos individuales de la manera que la neurociencia animal invasiva puede hacerlo. Por ejemplo, la amígdala del primate no humano se puede dividir en 13 núcleos y áreas corticales (Freese & Amaral, 2009). Estas regiones de la amígdala realizan diferentes funciones. El núcleo central envía salidas que involucran áreas del tronco encefálico que dan como resultado expresiones emocionales innatas y respuestas fisiológicas asociadas. El núcleo basal está conectado con áreas estriatales que están involucradas con acciones como correr hacia la seguridad. Además, no es posible hacer mapas uno a uno de las emociones en las regiones cerebrales. Por ejemplo, una extensa investigación ha examinado la implicación de la amígdala en el miedo, pero la investigación también ha demostrado que la amígdala es activa durante la incertidumbre (Whalen, 1998) así como emociones positivas (Anderson et al., 2003; Schulkin, 1990).
Conclusión
La investigación en neurociencia afectiva ha contribuido al conocimiento sobre los procesos emocionales, motivacionales y conductuales. El estudio de los sistemas emocionales básicos de animales no humanos proporciona información sobre la organización y desarrollo de emociones humanas más complejas. Aunque aún queda mucho por descubrir, los hallazgos actuales en la neurociencia afectiva ya han influido en nuestra comprensión del consumo y abuso de drogas, trastornos psicológicos como el trastorno de pánico, y emociones humanas complejas como el deseo y el disfrute, el dolor y el amor.
Recursos Externos
- Video: Entrevista de 1 hora a Jaak Panksepp, el padre de la neurociencia afectiva
- Video: Una entrevista de 15 minutos con Kent Berridge sobre el placer en el cerebro
- Video: Entrevista de 5 minutos con Joseph LeDoux sobre la amígdala y el miedo
- Web: Modelo 3D interactivo de anatomía cerebral
- http://www.pbs.org/wnet/brain/3d/index.html
Preguntas de Discusión
- Los circuitos neuronales de “gusto” son diferentes de los circuitos de “querer”. ¿Cómo podría relacionarse esto con los problemas que encuentran las personas cuando hacen dieta, combaten las adicciones o intentan cambiar otros hábitos?
- Las estructuras y neurotransmisores que producen placer durante el contacto social también producen pánico y dolor cuando los organismos se ven privados de contacto social. ¿Cómo contribuye esto a una comprensión del amor?
- La investigación muestra que los ambientes estresantes aumentan el área del núcleo accumbens que es sensible al miedo, mientras que los ambientes preferidos aumentan el área que es sensible a las recompensas. ¿Cómo podrían ser adaptativos estos cambios?
El vocabulario
- Afectar
- Un proceso emocional; incluye estados de ánimo, sentimientos subjetivos y emociones discretas.
- Amígdala
- Dos estructuras almendradas localizadas en los lóbulos temporales mediales del cerebro.
- Hipotálamo
- Una estructura cerebral ubicada debajo del tálamo y por encima del tronco encefálico.
- Neurociencia
- El estudio del sistema nervioso.
- Núcleo accumbens
- Una región del prosencéfalo basal ubicada frente a la región preóptica.
- Corteza Orbital Frontal
- Una región de los lóbulos frontales del cerebro por encima de las cuencas oculares.
- Gris periacueductal
- La materia gris en el mesencéfalo cerca del acueducto cerebral.
- Región preóptica
- Una parte del hipotálamo anterior.
- Estria terminalis
- Una banda de fibras que discurre a lo largo de la superficie superior del tálamo.
- Tálamo
- Una estructura en la línea media del cerebro ubicada entre el mesencéfalo y la corteza cerebral.
- Corteza visual
- La parte del cerebro que procesa la información visual, ubicada en la parte posterior del cerebro.
Referencias
- Anderson, A. K., Christoff, K., Stappen, I., Panitz, D., Ghahremani, D. G., Glover, G.,. Sobel, N. (2003). Representaciones neuronales disociadas de intensidad y valencia en el olfato humano. Neurociencia de la naturaleza, 6, 196—202.
- Berkman, E. T., & Lieberman, M. D. (2010). Acercarse a lo malo y evitar lo bueno: La asimetría cortical prefrontal lateral distingue entre acción y valencia. Revista de Neurociencia Cognitiva, 22 (9), 1970—1979. doi: 10.1162/jocn.2009.21317
- Berridge, K. C., & Kringelbach, M. L. (2013). Neurociencia del afecto: mecanismos cerebrales de placer y desagrado. Dictamen Actual en Neurobiología, 23, 294—303. doi.org/10.1016/j.conb.2013.01.017
- Berridge, K. C., & Kringelbach, M. L. (2008). Neurociencia afectiva del placer: Recompensa en humanos y animales. Psicofarmacología, 199, 457—480. doi: 10.1007/s00213-008-1099-6
- Blanchard, D. C., & Blanchard, R. J. (2003). ¿Qué nos puede decir la investigación sobre agresión animal sobre la agresión humana? Hormonas y Conducta, 44, 171—177.
- Farb, N.A.S., Chapman, H. A., & Anderson, A. K. (2013). Emociones: La forma sigue a la función. Dictamen Actual en Neurobiología, 23, 393—398. http://dx.doi.org/10.1016/j.conb.2013.01.015
- Fox, N. A., & Davidson, R. J. (1986). Cambios gustativos en los signos faciales de emoción y la asimetría de la actividad eléctrica cerebral en recién nacidos humanos. Neuropsicologia, 24, 417—422.
- Freese, J. L., & Amaral, D. G. (2009). Neuroanatomía de la amígdala de primates. En P. J. Whalen & E. A. Phelps (Eds.), La amígdala humana (pp. 3—42). Nueva York, NY: Guilford Press.
- Gable, P. A., & Harmon-Jones, E. (2008). Activación frontal izquierda relativa a estímulos apetitivos: Considerando el papel de las diferencias individuales. Psicofisiología, 45, 275-278.
- Goldstein, K. (1939). El organismo: Un enfoque holístico de la biología, derivado de datos patológicos en el hombre. Nueva York, NY: Libro Americano.
- Gottfried, J. A., O'Doherty, J., & Dolan, R. J. (2002). Aprendizaje olfativo apetitivo y aversivo en humanos estudiados mediante resonancia magnética funcional relacionada con eventos. Revista de Neurociencia, 22, 10829—10837.
- Gray, J. A. (1987). La psicología del miedo y el estrés (2ª ed.). Cambridge, Inglaterra: Cambridge University Press.
- Harmon-Jones, E., Harmon-Jones, C., & Price, T. F. (2013). ¿Qué es la motivación de aproximación? Emotion Review, 5, 291—295. doi: 10.1177/1754073913477509
- Heinrichs, M., von Dawans, B., & Domes, G. (2009). Oxitocina, vasopresina y comportamiento social humano. Fronteras en Neuroendocrinología, 30, 548—557.
- Izard, C. E. (2010). Los muchos significados/aspectos de la emoción: Definiciones, funciones, activación y regulación. Emotion Review, 2, 363—370. doi: 10.1177/1754073910374661
- LeDoux, J. E. (1996). El cerebro emocional: Los misteriosos fundamentos de la vida emocional. Nueva York, NY: Simon & Schuster.
- LeDoux, J. E., Farb, C. F., Ruggiero, D. A. (1990). Organización topográfica de neuronas en el tálamo acústico que se proyectan hacia la amígdala. Revista de Neurociencia, 10, 1043—1054.
- Numan, M., & Insel, T. R. (2003). La neurobiología del comportamiento parental. Nueva York, NY: SpringerVerlag.
- O'Doherty J. P., Deichmann, R., Critchley, H. D., & Dolan, R. J. (2002). Respuestas neuronales durante la anticipación de una recompensa gustativa primaria. Neurona, 33, 815—826.
- O'Doherty, J., Kringelbach, M. L., Rolls, E. T., Hornak, J., & Andrews, C. (2001). Representaciones abstractas de recompensa y castigo en la corteza orbitofrontal humana. Neurociencia de la naturaleza, 4, 95—102.
- O'Doherty, J., Winston, J., Critchley, H., Perrett, D., Burt, D. M., & Dolan, R. J. (2003). Belleza en una sonrisa: El papel de la corteza orbitofrontal medial en el atractivo facial. Neuropsicologia, 41, 147—155.
- Panksepp, J. (1998). Neurociencia afectiva: Los fundamentos de las emociones humanas y animales. Nueva York, NY: Oxford University Press.
- Panksepp, J., & Biven, L. (2012). La arqueología de la mente: Orígenes neuroevolutivos de las emociones humanas. Nueva York, NY: Norton.
- Panksepp, J., Normansell, L., Herman, B., Bishop, P., & Crepeau, L. (1988). Control neuronal y neuroquímico de la llamada de socorro de separación. En J. D. Newman (Ed.), El control fisiológico de la vocalización de mamíferos (pp. 263—299). Nueva York, NY: Pleno.
- Peterson, C. K., & Harmon-Jones, E. (2012). Ira y testosterona: Evidencia de que la ira inducida situacionalmente se relaciona con la testosterona inducida situacionalmente. Emoción, 12, 899—902. doi: 10.1037/a0025300
- Reynolds, S. M., & Berridge, K. C. (2008). Ambientes emocionales reintonizan la valencia de las funciones apetitivas versus temerosas en el núcleo accumbens. Neurociencia de la naturaleza, 11, 423—425.
- Reynolds, S. M., & Berridge, K. C. (2002). Motivación positiva y negativa en la cáscara del núcleo accumbens: Gradientes rostrocaudales bivalentes para comer provocado por GABA, reacciones de “gusto”/“disgusto”, preferencia de lugar/evitación y miedo. Revista de Neurociencia, 22, 7308—7320.
- Reynolds, S. M., & Berridge, K. C. (2001). Miedo y alimentación en la cáscara del núcleo accumbens: segregación rostrocaudal del comportamiento defensivo provocado por GABA versus comportamiento alimentario. Revista de Neurociencia, 21, 3261—3270.
- Schulkin, J. (1991). Hambre de sodio: La búsqueda de un sabor salado. Nueva York, NY: Cambridge University Press.
- Seymour, B., Daw, N., Dayan, P., Singer, T., & Dolan, R. (2007). Codificación diferencial de pérdidas y ganancias en el cuerpo estriado humano. Revista de Neurociencia, 27, 4826—4831.
- Stewart, J., De Wit, H., & Eikelboom, R. (1984). Papel de los efectos no condicionados y condicionados de drogas en la autoadministración de opiáceos y estimulantes. Revisión Psicológica, 91, 251-268.
- Wacker, J., Mueller, E. M., Pizzagalli, D. A., Hennig, J., & Stemmler, G. (2013). El bloqueo de dopamina-receptor D2 invierte la asociación entre la motivación del enfoque rasgo y la asimetría frontal en un contexto enfoque-motivación. Ciencia Psicológica, 24 (4), 489—497. doi: 10.1177/0956797612458935
- Whalen, P. J. (1998). Miedo, vigilancia y ambigüedad: estudios iniciales de neuroimagen de la amígdala humana. Direcciones Actuales en Ciencia Psicológica, 7, 177—188.