
8.6: El Sistema Vestibular
- Page ID
- 144010

Colegio de Medicina de Baylor
El sistema vestibular funciona para detectar el movimiento y la posición de la cabeza en relación con la gravedad y está involucrado principalmente en el control fino de la mirada visual, la postura, la ortostasia, la orientación espacial y la navegación. Las señales vestibulares están altamente procesadas en muchas regiones del cerebro y están involucradas en muchas funciones esenciales. En este módulo, proporcionamos una visión general de cómo funciona el sistema vestibular y cómo se utilizan las señales vestibulares para guiar el comportamiento.
objetivos de aprendizaje
- Definir las estructuras básicas del sistema receptor vestibular.
- Describir la neuroanatomía de las vías vestibuloocular, vestibuloespinal y vestibulo-talamo-corticales.
- Describir el sistema comisural vestibular.
- Describir las diferentes áreas corticales multisensoriales para la percepción del movimiento.
Introducción
¿Recuerdas la sensación de mareo que tenías cuando eras niño después de saltar del tiovivo o dar vueltas como un top? Estos sentimientos resultan de la activación del sistema vestibular, que detecta nuestros movimientos a través del espacio pero no es un sentido consciente como la visión o el oído. De hecho, la mayoría de las funciones vestibulares son imperceptibles, pero las sensaciones relacionadas con los vestibulares, como el mareo por movimiento, pueden aparecer rápidamente al montar en una montaña rusa, tener un viaje en avión lleno de baches o navegar en un barco en mares agitados. Sin embargo, estas sensaciones son realmente efectos secundarios y el sistema vestibular es realmente extremadamente importante para las actividades cotidianas, ya que las señales vestibulares están involucradas en gran parte del procesamiento de la información del cerebro que controla funciones tan fundamentales como el equilibrio, la postura, la estabilización de la mirada, el espacio orientación, y navegación, por nombrar algunos. En muchas regiones del cerebro, la información vestibular se combina con señales de los otros sentidos así como con información motora para dar lugar a la percepción del movimiento, la conciencia corporal y el control conductual. Aquí, exploraremos el funcionamiento del sistema vestibular y consideraremos algunos de los cálculos integrados que realiza el cerebro utilizando señales vestibulares para guiar nuestro comportamiento común.

Estructura de los receptores vestibulares
Los receptores vestibulares se encuentran en el oído interno junto a la cóclea auditiva. Detectan movimiento rotacional (giros de cabeza), movimiento lineal (traducciones) e inclinaciones de la cabeza en relación con la gravedad y transducen estos movimientos en señales neuronales que pueden enviarse al cerebro. Hay cinco receptores vestibulares en cada oído (módulo de audición, Figura 8.6.1- http://noba.to/jry3cu78), incluyendo tres canales semicirculares (horizontal, anterior y posterior) que transducen aceleraciones angulares rotacionales y dos receptores de otolitos (utrículo y saccule) que transduzcan aceleraciones lineales (Lindeman, 1969). Juntos, los canales semicirculares y los órganos del otolito pueden responder al movimiento de la cabeza y mantener la posición estática de la cabeza en relación con la gravedad en todas las direcciones en el espacio 3D.
Estos receptores están contenidos en una serie de tubos rellenos de fluido interconectados que están protegidos por un hueso denso y suprayacente (Iurato, 1967). Cada uno de los tres canales semicirculares se encuentra en un plano que es ortogonal a los otros dos. El canal semicircular horizontal se encuentra en un plano de cabeza aproximadamente horizontal, mientras que los canales semicirculares anterior y posterior se encuentran verticalmente en la cabeza (Blanks, Curthoys, Bennett, & Markham, 1985). Las células receptoras del canal semicircular, denominadas células ciliadas, se localizan solo en el medio de los tubos circulares en un epitelio especial, cubierto por una membrana gelatinosa que se extiende a través del tubo para formar un sello hermético a los fluidos como la piel de un tambor (Figuras 1A y 1B). Las células ciliadas se llaman así debido a una matriz de casi 100 estereocilia de altura escalonada (como un órgano de tubo de iglesia) que sobresalen de la parte superior de la célula hacia la membrana de gelatina suprayacente (Wersäll, 1956). Las estereocilia más cortas están en un extremo de la célula y las más altas en el otro (Lindeman, 1969). Cuando se gira la cabeza, el fluido en los canales semicirculares se queda atrás del movimiento de la cabeza y empuja sobre la membrana de gelatina, que dobla la estereocilia.

Como se muestra en la Figura 8.6.2, cuando la cabeza se mueve hacia las células ciliadas receptoras (por ejemplo, la cabeza izquierda gira para el canal semicircular horizontal izquierdo), los estereocilios se doblan hacia el extremo más alto y se abren canales iónicos especiales mecánicamente cerrados en las puntas de los cilios, lo que excita ( despolariza) la célula (Shotwell, Jacobs, & Hudspeth, 1981). El movimiento de la cabeza en la dirección opuesta provoca la flexión hacia la estereocilia más pequeña, lo que cierra los canales e inhibe (hiperpolariza) la célula. Los canales semicirculares del oído izquierdo y derecho tienen polaridad opuesta, así por ejemplo, cuando giras la cabeza hacia la izquierda, los receptores en el canal semicircular horizontal izquierdo se excitarán mientras que los receptores del canal horizontal del oído derecho serán inhibidos (Figura 8.6.3). La misma relación es cierta para los canales semicirculares verticales. Las fibras nerviosas aferentes vestibulares inervan la base de la célula pilosa y aumentan o disminuyen su velocidad de disparo neural a medida que la célula receptora se excita o inhibe (Dickman y Correia, 1989), respectivamente, y luego llevan estas señales con respecto al movimiento rotacional de la cabeza al cerebro como parte de la Nervio vestibulococlear (Nervio craneal VIII). Entran en el tronco encefálico y terminan en los núcleos vestibulares ipsilaterales, el cerebelo y la formación reticular (Carleton & Carpenter, 1984; Dickman & Fang, 1996). La célula pilosa vestibular primaria y los neurotransmisores aferentes son glutamato y aspartato. Debido a las propiedades mecánicas del sistema receptor vestibular, las aceleraciones rotacionales de la cabeza se integran en señales de velocidad (Van Egmond, Groen, & Jongkess, 1949) que luego son codificadas por aferentes de canal semicircular (Fernandez & Goldberg, 1971). Los umbrales de detección para el movimiento de rotación han demostrado que los aferentes pueden discriminar diferencias en la velocidad de la cabeza del orden de 2 grados/seg, pero también son sensibles a una amplia gama de movimientos naturales de la cabeza hasta altas velocidades de cabeza en los cientos de grados/seg (como puede experimentar cuando realiza una giro rápido de la cabeza hacia un sonido fuerte, o están realizando gimnasia; Sadeghi, Chacron, Taylor, & Cullen, 2007; Yu, Dickman, & Angelaki, 2012).
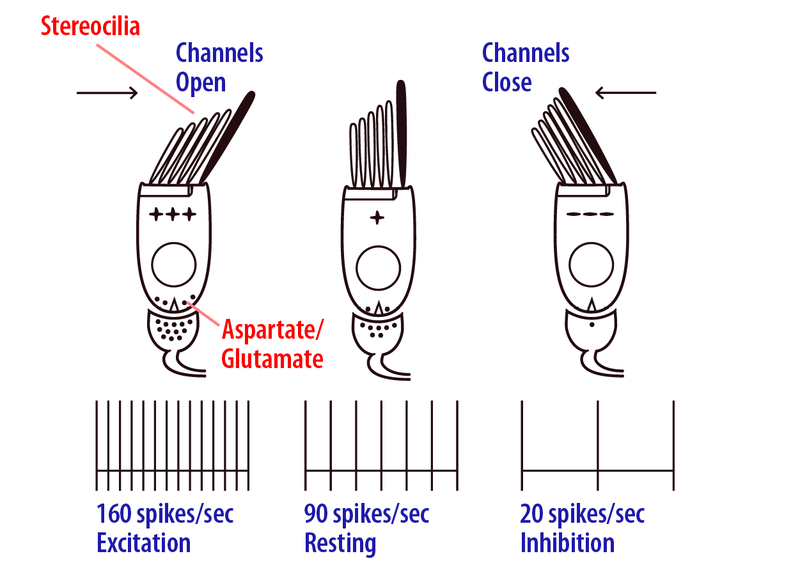

Los receptores de otolitos son sensibles a aceleraciones lineales e inclinaciones de la cabeza en relación con la gravedad (Fernandez & Goldberg, 1976a). El receptor de otolitos de utrículo se encuentra paralelo al canal semicircular horizontal y el receptor sáculo se encuentra vertical en la cabeza (Módulo de audición, Figura 8.6.1- http://noba.to/jry3cu78). Como se muestra en la Figura 8.6.4, un epitelio de otolito especial contiene células ciliadas receptoras cuya estereocilia se extiende dentro de una membrana de gelatina que está cubierta por una capa de cristales de carbonato de calcio, denominada otoconia, como rocas apiladas para formar un embarcadero (Lindeman, 1969). La otoconia no se ve afectada por movimientos fluidos, sino que son desplazadas por aceleraciones lineales, incluyendo traslaciones (por ejemplo, movimientos hacia adelante/atrás o hacia arriba/hacia abajo) o cambios en la posición de la cabeza en relación con la gravedad. Estas aceleraciones lineales producen desplazamientos de la otoconia (debido a su gran masa), al igual que rocas rodando cuesta abajo o tu taza de café cayendo del salpicadero del auto cuando empujas el acelerador. Los movimientos de la otoconia doblan la estereocilia de las células ciliadas y los canales de apertura/cierre de manera similar a la descrita para los canales semicirculares. Sin embargo, las células ciliadas del otolito están polarizadas de tal manera que las estereocilia más altas apuntan hacia el centro del utrículo y alejadas del centro en el sáculo, lo que efectivamente divide los receptores en dos grupos opuestos (Flock, 1964; Lindeman, 1969). De esta manera, algunas células ciliadas se excitan y algunas se inhiben por cada fuerza de movimiento lineal o inclinación de la cabeza experimentada, con la población de receptores y sus aferentes inervantes sintonizados direccionalmente a todos los movimientos o inclinaciones de la cabeza en el espacio 3D (Fernandez & Goldberg, 1976b).

Todas las células ciliadas vestibulares y aferentes reciben conexiones de eferentes vestibulares, que son fibras que se proyectan desde el cerebro hacia fuera a los órganos receptores vestibulares, cuya función no se entiende bien. Se piensa que los eferentes controlan la sensibilidad del receptor (Boyle, Carey, & Highstein, 1991). El neurotransmisor eferente primario es la acetilcolina (Anniko & Arnold, 1991).
Núcleos vestibulares
Los núcleos vestibulares comprenden un gran conjunto de elementos neurales en el tronco encefálico que reciben movimiento y otras señales multisensoriales, luego regulan las respuestas de movimiento y la experiencia sensorial. Muchas neuronas de núcleos vestibulares tienen conexiones recíprocas con el cerebelo que forman importantes mecanismos reguladores para el control de los movimientos oculares, los movimientos de la cabeza y la postura. Hay cuatro núcleos vestibulares principales que se encuentran en la médula rostral y los pones caudales del tronco encefálico; todos reciben entrada directa de aferentes vestibulares (Brodal, 1984; Precht y Shimazu, 1965). Muchas de estas neuronas de núcleos reciben información de movimiento convergente del oído opuesto a través de una vía comisural inhibitoria que utiliza ácido gamma-aminobutírico (GABA) como neurotransmisor (Kasahara y Uchino, 1974; Shimazu y Precht, 1966). La vía comisural está altamente organizada de tal manera que las células que reciben señales del canal excitatorio horizontal del oído ipsilateral también recibirán señales de canal horizontal inhibitorias contralaterales del oído opuesto. Este hecho da lugar a una función vestibular “push-pull”, por lo que la sensibilidad direccional a el movimiento de la cabeza está codificado por señales opuestas del receptor. Debido a que las neuronas de los núcleos vestibulares reciben información de receptores bilaterales del oído interno y debido a que mantienen una alta tasa de disparo espontáneo (casi 100 impulsos/seg), se cree que actúan para “comparar” las tasas de descarga relativa de la actividad de disparo aferente del canal izquierdo vs derecho. Por ejemplo, durante un giro hacia la izquierda de la cabeza, las neuronas de los núcleos del tronco encefálico izquierdo reciben información de alta tasa de activación del canal horizontal izquierdo e información de baja tasa de activación del canal horizontal derecho. La comparación de actividad se interpreta como un giro de cabeza izquierda. Las respuestas neuronales de núcleos similares existen cuando la cabeza está inclinada o enrollada, siendo los canales semicirculares verticales estimulados por el movimiento rotacional en sus planos de sensibilidad. Sin embargo, la respuesta de vaivén opuesta de los canales verticales ocurre con el canal semicircular anterior en una oreja y el canal semicircular posterior coplanario de la oreja opuesta. El daño o enfermedad que interrumpe la información de la señal del oído interno desde un lado de la cabeza puede cambiar la actividad normal de reposo en las fibras aferentes del nervio VIII y será interpretada por el cerebro como una rotación de la cabeza, aunque la cabeza esté estacionaria. Estos efectos a menudo conducen a ilusiones de girar o rotar que pueden ser bastante molestas y pueden producir náuseas o vómitos. Sin embargo, con el tiempo las fibras comisurales proporcionan compensación vestibular, proceso por el cual la pérdida de la función del receptor vestibular unilateral se restablece parcialmente centralmente y las respuestas conductuales, como el reflejo vestibuloocular (VOR) y las respuestas posturales, principalmente recuperar (Beraneck et al., 2003; Fetter & Zee, 1988; Newlands, Hesse, Haque, & Angelaki, 2001; Newlands y Perachio, 1990).
Además de la vía comisural, muchas neuronas de núcleos vestibulares reciben señales propioceptivas de la médula espinal con respecto al movimiento y posición muscular, señales visuales con respecto al movimiento espacial, otras señales multisensoriales (por ejemplo, trigémino) y señales de orden superior de la corteza. Se piensa que las entradas corticales regulan la mirada fina y el control postural, así como suprimen los reflejos compensatorios normales durante el movimiento para provocar movimientos volitivos. De especial importancia son las señales convergentes del canal semicircular y aferentes del otolito que permiten a las neuronas vestibulares centrales calcular propiedades específicas del movimiento de la cabeza (Dickman & Angelaki, 2002). Por ejemplo, Einstein (1907) demostró que las aceleraciones lineales son equivalentes tanto si surgen del movimiento de traslación como de las inclinaciones de la cabeza en relación con la gravedad. Los receptores de otolitos no pueden discriminar entre los dos, entonces, ¿cómo es que podemos notar la diferencia entre cuando estamos traduciendo hacia adelante e inclinándonos hacia atrás, donde la aceleración lineal señalada por los aferentes otolitos es la misma? Los núcleos vestibulares y las neuronas cerebelosas utilizan señales convergentes tanto de los canales semicirculares como de los receptores de otolitos para discriminar entre inclinación y traducción, y como resultado, algunas células codifican la inclinación de la cabeza (Zhou, 2006) mientras que otras células codifican el movimiento traslacional (Angelaki, Shaikh, Green, & Dickman, 2004).
Sistema vestibuloocular
El sistema vestibular es responsable de controlar la estabilidad de la mirada durante el movimiento (Crane & Demer, 1997). Por ejemplo, si queremos leer el letrero en el escaparate de una tienda mientras caminamos, debemos mantener la fijación foveal en las palabras mientras compensamos los movimientos combinados de rotación y traslación de la cabeza incurridos durante nuestro paso. El sistema vestibular regula los movimientos compensatorios de ojos, cuello, espinales y extremidades para mantener la mirada (Keshner & Peterson, 1995). Uno de los principales componentes que contribuyen a la estabilidad de la mirada es el VOR, que produce movimientos oculares reflexivos que son iguales en magnitud y opuestos en dirección al movimiento percibido de la cabeza en el espacio 3D (Wilson et al., 1995). El VOR es tan preciso y rápido que permite a las personas mantener la fijación visual sobre los objetos de interés mientras experimentan condiciones de movimiento exigentes, como correr, esquiar, jugar al tenis y conducir. De hecho, se ha demostrado que la estabilización de la mirada en humanos es completamente compensatoria (esencialmente perfecta) para la mayoría de los comportamientos naturales. Para producir el VOR, las neuronas vestibulares deben controlar cada uno de los seis pares de músculos oculares al unísono a través de un conjunto específico de conexiones con los núcleos oculomotores (Ezure & Graf, 1984). Los canales semicirculares anterior y posterior junto con el sáculo controlan los movimientos oculares verticales y torsionales (giro del ojo alrededor de la línea de visión), mientras que los canales horizontales y el utrículo controlan los movimientos oculares horizontales.
Para entender cómo funciona el VOR, tomemos el ejemplo de la respuesta compensatoria para un giro de cabeza hacia la izquierda mientras leemos las palabras en la pantalla de una computadora. La vía básica consiste en aferentes de canal semicircular horizontal que se proyectan hacia neuronas específicas en los núcleos vestibulares. Estas células nucleicas, a su vez, envían una señal excitatoria al núcleo abducens contralateral, el cual se proyecta a través del sexto nervio craneal para inervar el músculo recto lateral (Figura 8.6.5). Algunas neuronas abducens envían una proyección excitatoria de vuelta a través de la línea media a una subdivisión de células en el núcleo oculomotor ipsilateral, que, a su vez, se proyecta a través del tercer nervio craneal para inervar el músculo recto medial derecho (ipsilateral). Cuando se realiza un giro de cabeza hacia la izquierda, los aferentes vestibulares del canal horizontal izquierdo aumentarán su velocidad de disparo y, en consecuencia, aumentarán la actividad de las neuronas de los núcleos vestibulares que se proyectan hacia el núcleo abducens derecho opuesto (contralateral). Las neuronas abducens producen contracción del recto lateral derecho y, a través de una proyección celular separada hacia el núcleo oculomotor izquierdo, excitan los músculos rectos mediales izquierdos. Además, las conexiones inhibitorias bilaterales coincidentes relajan los músculos rectos laterales izquierdo y recto medial derecho del ojo. El movimiento resultante del ojo hacia la derecha para ambos ojos estabiliza el objeto de interés sobre la retina para obtener la mayor agudeza visual.
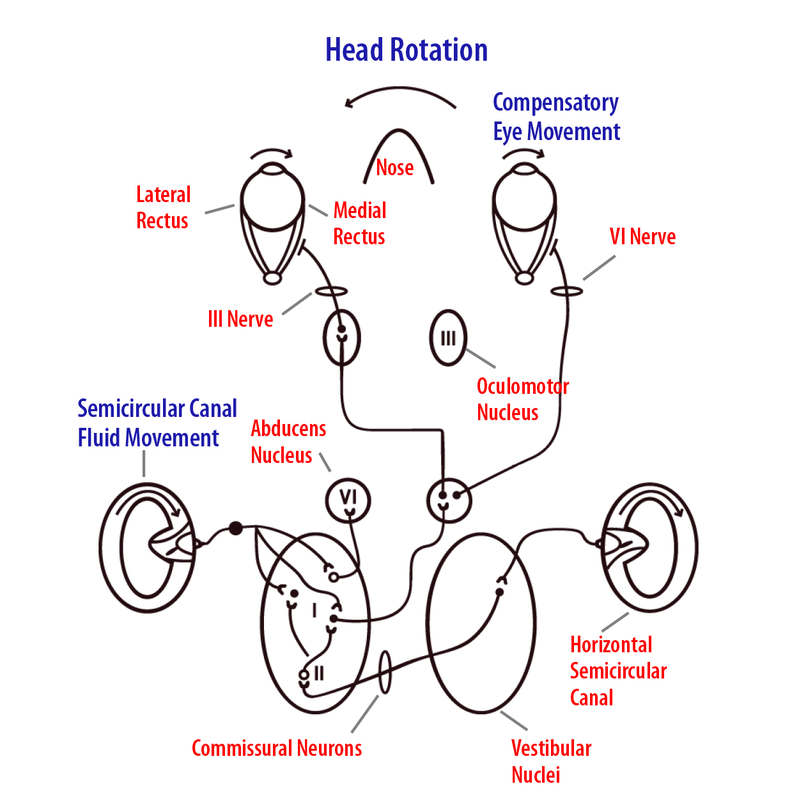
Durante las traducciones lineales, también ocurre un tipo diferente de VOR (Paige & Tomko, 1991). Por ejemplo, el movimiento lateral hacia la izquierda da como resultado un movimiento horizontal del ojo hacia la derecha para mantener la estabilidad visual en un objeto de interés. De manera similar, los movimientos verticales de arriba a abajo de la cabeza (como ocurren al caminar o correr) provocan movimientos oculares verticales dirigidos opuestamente (Angelaki, McHenry, & Hess, 2000). Para estos reflejos, la amplitud del VOR traslacional depende de la distancia de visión. Esto se debe a que el ángulo de vergencia (es decir, el ángulo entre las líneas de visión para cada ojo) varía en función de la inversa de la distancia al objeto visual visto (Schwarz, Busettini, & Miles, 1989). Los objetos visuales que están muy lejos (2 metros o más) no requieren ángulo de vergencia, pero a medida que los objetos visuales se acercan (por ejemplo, al sostener el dedo cerca de la nariz), se necesita un ángulo de vergencia grande. Durante el movimiento de traslación, los ojos cambiarán su ángulo de vergencia a medida que el objeto visual se mueve de cerca a más lejos (o viceversa). Estas respuestas son resultado de la activación de los receptores otolitos, con conexiones a los núcleos oculomotores similares a las descritas anteriormente para el reflejo vestibuloocular rotacional. Con las inclinaciones de la cabeza, el movimiento ocular resultante se denomina torsión, y consiste en un movimiento rotacional del ojo alrededor de la línea de visión que está en la dirección opuesta a la inclinación de la cabeza. Como se mencionó anteriormente, existen importantes conexiones recíprocas entre los núcleos vestibulares y el cerebelo. Está bien establecido que estas conexiones son cruciales para el aprendizaje motor adaptativo en el reflejo vestibuloocular (Lisberger, Pavelko, & Broussard, 1994).
Red vestibulo-espinal

Existen dos vías descendentes vestibulares que regulan las respuestas musculares corporales al movimiento y la gravedad, consistentes en el tracto vestibulo-espinal lateral (LVST) y el tracto vestibulo-espinal medial (MVST). El control reflexivo de los músculos de cabeza y cuello surge a través de las neuronas en el tracto vestibuloespinal medial (MVST). Estas neuronas comprenden el reflejo vestibulocólico rápido (VCR) que sirve para estabilizar la cabeza en el espacio y participa en el control de la mirada (Peterson, Goldber, Bilotto, & Fuller, 1985). Las neuronas MVST reciben entrada de los receptores vestibulares y del cerebelo, e información somatosensorial de la médula espinal. Las neuronas MVST transportan señales excitadoras e inhibidoras para inervar las neuronas motoras flexoras y extensoras del cuello en la médula espinal. Por ejemplo, si uno tropieza sobre una grieta en el pavimento mientras camina, las neuronas MVST recibirán señales de aceleración lineal hacia abajo y hacia adelante de los receptores de otolitos y señales de aceleración de rotación hacia adelante de los canales semicirculares verticales. El VCR compensará proporcionando señales excitatorias a los músculos flexores del cuello dorsal y señales inhibitorias a los músculos extensores ventrales del cuello, que mueve la cabeza hacia arriba y opuesta al movimiento de caída para protegerla del impacto.
El LVST comprende una organización topográfica de células de núcleos vestibulares que reciben entrada sustancial del cerebelo, entradas propioceptivas de la médula espinal y señales aferentes convergentes de receptores vestibulares. Las fibras LVST se proyectan ipsilaterales a muchos niveles de neuronas motoras en el cordón para proporcionar coordinación de diferentes grupos musculares para el control postural (Shinoda, Sugiuchi, Futami, Ando, & Kawasaki, 1994). Las neuronas LVST contienen acetilcolina o glutamato como neurotransmisor y ejercen una influencia excitadora sobre las neuronas motoras del músculo extensor. Por ejemplo, las fibras LVST producen extensión de la musculatura axial contralateral y de las extremidades cuando el cuerpo está inclinado lateralmente. Estas acciones sirven para estabilizar el centro de gravedad del cuerpo con el fin de preservar la postura erguida.
Control vestibulo-autonómico
Algunas neuronas del núcleo vestibular envían proyecciones a la formación reticular, a los núcleos pontinos dorsales y al núcleo del tracto solitario. Estas conexiones regulan la respiración y la circulación a través de respuestas autónomas vestibulares compensatorias que estabilizan la respiración y la presión arterial durante el movimiento corporal y los cambios relativos a la gravedad. También pueden ser importantes para la inducción del mareo por movimiento y la emesis.
Señales vestibulares en el tálamo y la corteza
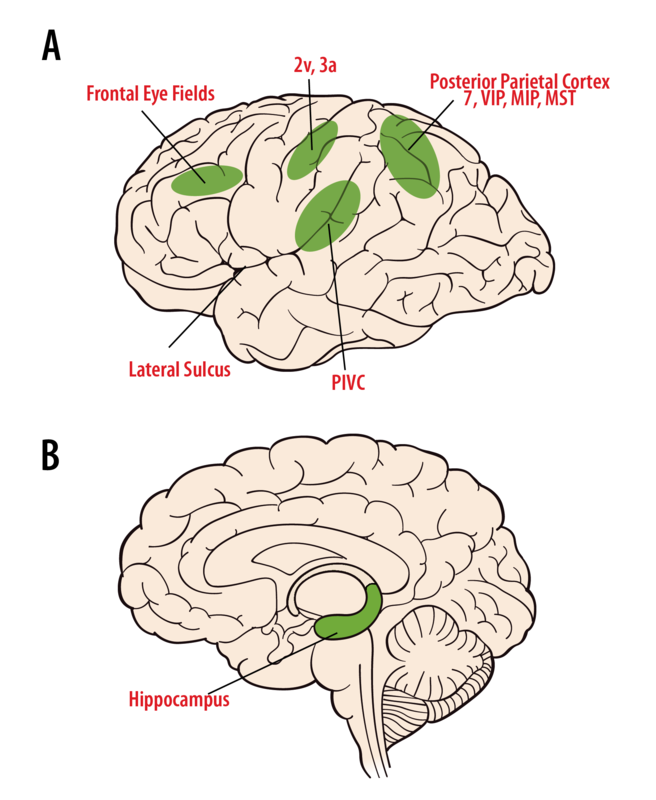
La percepción cognitiva del movimiento, la orientación espacial y la navegación a través del espacio surge a través de información multisensorial a partir de señales vestibulares, visuales y somatosensoriales en el tálamo y la corteza (Figura 8.6.6). Las neuronas de los núcleos vestibulares se proyectan bilateralmente a las diversas regiones talámicas. Las neuronas en el grupo ventral posterior responden a señales vestibulares solas, o a señales vestibulares más somatosensoriales, y se proyectan a la corteza somatosensorial primaria (área 3a, 2v), corteza de asociación somatosensorial, corteza parietal posterior (áreas 5 y 7) y la insula de la corteza temporal (Marlinski & McCrea, 2008; Meng, May, Dickman, & Angelaki, 2007). El grupo nuclear posterior (PO), cerca del cuerpo geniculado medial, recibe señales vestibulares y auditivas, así como entradas del colículo superior y la médula espinal, lo que indica una integración de múltiples señales sensoriales. Algunas neuronas pulvinares anteriores también responden a estímulos de movimiento y se proyectan hacia el área cortical 3a, la insula posterior y la corteza temporo-parietal (PIVC). En los humanos, la estimulación eléctrica de las zonas talámicas produce sensaciones de movimiento y a veces mareos.
Las células del área 2v responden al movimiento, y la estimulación eléctrica de esta área en humanos produce sensaciones de movimiento, giro o mareos. El área 3a se encuentra en la base del surco central adyacente a la corteza motora y se cree que está involucrada en el control motor integrador de la cabeza y el cuerpo (Guldin, Akbarian, & Grusser, 1992). Las neuronas en el PIVC son multisensoriales, responden al movimiento corporal, somatosensoriales, propioceptivos y estímulos de movimiento visual (Chen, DeAngelis, & Angelaki, 2011; Grusser, Pause, & Schreiter, 1982). PIVC y las áreas 3a y 2v están fuertemente interconectadas. También se han observado neuronas vestibulares en la corteza parietal posterior; en el área 7, en el área intraparietal ventral (VIP), el área intraparietal medial (MIP) y el área temporal media superior (MST). VIP contiene neuronas multimodales involucradas en la codificación espacial. Las neuronas MIP y MST responden al movimiento corporal a través del espacio mediante la integración multisensorial del movimiento visual y las señales vestibulares (Gu, DeAngelis, & Angelaki, 2007) y muchas células MST están directamente involucradas en la percepción de rumbo (Gu, Watkins, Angelaki y DeAngelis, 2006). Las lesiones de las áreas corticales parietales pueden dar lugar a confusiones en la conciencia espacial. Finalmente, las áreas involucradas con el control de las sacadas y los movimientos oculares de persecución, incluyendo el área 6, el área 8 y la circunvolución frontal superior, reciben señales vestibulares (Fukushima, Sato, Fukushima, Shinmei y Kaneko, 2000). Todavía no se comprende bien cómo estas diferentes regiones corticales contribuyen a nuestra percepción del movimiento y la orientación espacial.
Orientación espacial y navegación
Nuestra capacidad para conocer dónde estamos y navegar por diferentes ubicaciones espaciales es esencial para la supervivencia. Se cree que un mapa cognitivo de nuestro entorno se crea a través de la exploración y luego se usa para orientación espacial y navegación, como conducir a la tienda, o caminar por una casa oscura (McNaughton, Battaglia, Jensen, Moser, & Moser, 2006). Se han identificado células en el sistema límbico y el hipocampo que contribuyen a estas funciones, incluyendo celdas de lugar, celdas de cuadrícula y celdas de dirección de cabeza (Figura 6B). Las células de lugar en el hipocampo codifican ubicaciones específicas en el ambiente (O'Keefe, 1976). Las células de cuadrícula en la corteza entorinal codifican mapas espaciales en un patrón teselado (Hafting, Fyhn, Molden, Moser, & Moser, 2005). Las células de dirección de la cabeza en el tálamo antero-dorsal codifican la dirección de rumbo, independientemente de la ubicación espacial (Taube, 1995). Se piensa que estos tipos de células trabajan juntos para proporcionar orientación espacial, memoria espacial y nuestra capacidad de navegar. Tanto las células de lugar como las células de dirección de la cabeza dependen de un sistema vestibular funcional para mantener su información direccional y de orientación (Stackman, Clark y Taube, 2002). El camino por el cual las señales vestibulares llegan a la red de navegación no se entiende bien; sin embargo, el daño al sistema vestibular, hipocampo y regiones del tálamo dorsal a menudo perturba nuestra capacidad de orientarnos en ambientes familiares, navegar de un lugar a otro, o incluso para encontrar nuestro camino a casa.
Meo por movimiento

Aunque una serie de condiciones pueden producir mareo por movimiento, generalmente se piensa que se evoca a partir de un desajuste en las señales sensoriales entre señales vestibulares, visuales y propioceptivas (Yates, Miller, & Lucot, 1998). Por ejemplo, leer un libro en un automóvil en una carretera sinuosa puede producir mareo por movimiento, por lo que las aceleraciones experimentadas por el sistema vestibular no coinciden con la entrada visual. Sin embargo, si uno mira por la ventana al paisaje que pasa durante el mismo viaje, no se produce ninguna enfermedad porque las señales visuales y vestibulares están alineadas. El mareo, una forma de mareo por movimiento, parece ser un caso especial y surge del inusual movimiento oscilatorio vertical y de balanceo. Estudios en humanos han encontrado que las oscilaciones de baja frecuencia de 0.2 Hz y las grandes amplitudes (como las que se encuentran en grandes mares durante una tormenta) tienen más probabilidades de causar mareo por movimiento, con frecuencias más altas que ofrecen pocos problemas.
Resumen
Aquí, hemos visto que el sistema vestibular transduce y codifica señales sobre el movimiento y la posición de la cabeza con respecto a la gravedad, información que luego es utilizada por el cerebro para muchas funciones y comportamientos esenciales. De hecho, entendemos mucho con respecto a las contribuciones vestibulares a los reflejos fundamentales, como los movimientos oculares compensatorios y el equilibrio durante el movimiento. Se ha avanzado más recientemente hacia la comprensión de cómo las señales vestibulares se combinan con otras señales sensoriales, como la visión, en el tálamo y la corteza para dar lugar a la percepción del movimiento. Sin embargo, existen muchas habilidades cognitivas complejas que sabemos que requieren información vestibular para funcionar, como la orientación espacial y los comportamientos de navegación, pero estos sistemas apenas están comenzando a ser investigados. Las investigaciones futuras sobre la función del sistema vestibular probablemente estarán orientadas a buscar respuestas a preguntas sobre cómo el cerebro hace frente a la pérdida de señal vestibular. De hecho, según los Institutos Nacionales de Salud, casi el 35% de los estadounidenses mayores de 40 años (69 millones de personas) han reportado problemas crónicos relacionados con los vestibulares. Por lo tanto, es de importancia significativa para la salud humana comprender mejor cómo las señales vestibulares contribuyen a las funciones cerebrales comunes y cómo se pueden realizar mejores opciones de tratamiento para la disfunción vestibular.
Recursos Externos
- Video Animado del Sistema Vestibular
- http://sites.sinauer.com/neuroscienc...ions14.01.html
Preguntas de Discusión
- Si una persona sufre pérdida de los receptores vestibulares en un oído debido a una enfermedad o trauma, ¿qué síntomas sufriría la persona? ¿Los síntomas serían permanentes?
- A menudo, el mareo por movimiento se alivia cuando una persona mira objetos a gran distancia, como cosas ubicadas en el horizonte lejano. ¿Por qué la visualización a distancia lejana ayuda en el mareo por movimiento mientras que la vista a distancia cercana (como leer un mapa o un libro) lo empeora
- Las señales vestibulares se combinan con señales visuales en ciertas áreas de la corteza y ayudan en la percepción del movimiento. ¿Qué tipos de señales proporciona el sistema visual para el automovimiento a través del espacio? ¿Qué tipos de señales vestibulares serían consistentes con el movimiento rotacional versus traslacional?
El vocabulario
- Núcleo abducens
- Un grupo de neuronas motoras excitatorias en el tronco encefálico medial que envían proyecciones a través del nervio craneal VITH para controlar el músculo recto lateral ipsilateral. Además, las interneuronas abducens envían una proyección excitatoria a través de la línea media a una subdivisión de células en el núcleo oculomotor ipsilateral, que se proyectan a través del nervio craneal IIIRD para inervar el músculo recto medial ipsilateral.
- Acetilcolina
- Un neurotransmisor compuesto orgánico que consiste en ácido acético y colina. Dependiendo del tipo de receptor, la acetilcolina puede tener efectos excitadores, inhibidores o moduladores.
- Fibras nerviosas aferentes
- Neuronas únicas que inervan las células ciliadas receptoras y transportan señales vestibulares al cerebro como parte del nervio vestibulococlear (nervio craneal VIII).
- Aspartato
- Un neurotransmisor de aminoácidos excitatorios que es ampliamente utilizado por los receptores vestibulares, aferentes y muchas neuronas en el cerebro.
- Reflejos compensatorios
- Reflejo motor estabilizador que ocurre en respuesta a un movimiento percibido, como el reflejo vestibuloocular, o las respuestas posturales que ocurren durante la carrera o el esquí.
- Despolarizado
- Cuando las células ciliadas receptoras tienen canales cerrados mecánicamente abiertos, la célula aumenta su voltaje de membrana, lo que produce una liberación de neurotransmisor para excitar la fibra nerviosa inervante.
- Umbrales de detección
- La menor cantidad de movimiento de la cabeza que puede ser reportada de manera confiable por un observador.
- Afinación direccional
- La dirección de movimiento preferida que exhiben las células ciliadas y aferentes donde se produce una respuesta excitatoria máxima y la dirección menos preferida donde no se produce respuesta. Se dice que las células están “sintonizadas” para una mejor y peor dirección de movimiento, con direcciones de movimiento intermedias que provocan una respuesta menor pero observable.
- Ácido gamma-aminobutírico
- Un neurotransmisor inhibidor importante en el sistema comisural vestibular.
- Estabilidad de la mirada
- Una combinación de respuestas de ojos, cuello y cabeza que están coordinadas para mantener la fijación visual (fóvea) sobre un punto de interés.
- Glutamato
- Un neurotransmisor de aminoácidos excitatorios que es ampliamente utilizado por los receptores vestibulares, aferentes y muchas neuronas en el cerebro.
- Células ciliadas
- Las células receptoras del sistema vestibular. Se les llama células ciliadas debido a los muchos cilios pilosos que se extienden desde la superficie apical de la célula hacia la membrana de gelatina. Los canales iónicos mecánicos en las puntas de los cilios se abren y cierran a medida que los cilios se doblan para provocar cambios en el voltaje de la membrana en la célula pilosa que son proporcionales a la intensidad y dirección del movimiento.
- Hiperpolariza
- Cuando las células ciliadas receptoras tienen canales cerrados mecánicamente, la célula disminuye su voltaje de membrana, lo que produce menos liberación de neurotransmisores para inhibir la fibra nerviosa inervante.
- Músculo recto lateral
- Un músculo ocular que gira hacia afuera en el plano horizontal.
- Tracto vestibulo-espinal lateral
- Neuronas vestibulares que se proyectan a todos los niveles de la médula espinal en el lado ipsilateral para controlar la postura y equilibrar los movimientos.
- Canales iónicos mecánicamente cerrados
- Canales iónicos localizados en las puntas de la estereocilia en las células receptoras que se abren y cierran a medida que los cilios se doblan hacia los cilios más altos/más pequeños, respectivamente. Estos canales son permeables a los iones de potasio, que abundan en el líquido bañando la parte superior de las células ciliadas.
- Tracto vestibulo-espinal medial
- Las neuronas del núcleo vestibular se proyectan bilateralmente a las neuronas motoras espinales cervicales para el control del movimiento de El tracto funciona principalmente en la dirección de la mirada y la estabilidad durante el movimiento.
- Neurotransmisores
- Compuesto químico utilizado para enviar señales de una célula receptora a una neurona, o de una neurona a otra. Los neurotransmisores pueden ser excitadores, inhibidores o moduladores y están empaquetados en pequeñas vesículas que se liberan desde los terminales terminales de las células.
- Núcleos oculomotores
- Incluye tres grupos neuronales en el tronco encefálico, el núcleo abducens, el núcleo oculomotor y el núcleo troclear, cuyas células envían órdenes motoras a los seis pares de músculos oculares.
- Núcleo oculomotor
- Un grupo de células en el tronco encefálico medio que contiene subgrupos de neuronas que se proyectan hacia los músculos recto medial, oblicuo inferior, recto inferior y recto superior de los ojos a través del 3er nervio craneal.
- Otoconia
- Pequeñas partículas de carbonato de calcio que se empaquetan en una capa en la parte superior de la membrana de gelatina que cubre la estereocilia de células ciliadas del receptor otolito.
- Receptores de otolitos
- Dos receptores vestibulares del oído interno (utrículo y sáculo) que transducen las aceleraciones lineales y la inclinación de la cabeza en relación con la gravedad en señales neuronales que luego se transfieren al cerebro.
- Proprioceptivo
- Información sensorial sobre la posición muscular y el movimiento derivado de receptores en los músculos, tendones y articulaciones.
- Canales semicirculares
- Un conjunto de tres receptores vestibulares del oído interno (horizontal, anterior, posterior) que transducen las aceleraciones rotacionales de la cabeza en señales de velocidad de rotación de la cabeza que luego se transfieren al cerebro. Hay tres canales semicirculares en cada oreja, siendo los planos principales de cada canal ortogonales entre sí.
- Estereocilia
- Proyecciones pilosas desde la parte superior de las células ciliadas receptoras. Las estereocilia se disponen en altura ascendente y cuando se desplazan hacia los cilios más altos, los canales mecánicos se abren y la célula se excita (despolariza). Cuando la estereocilia se desplaza hacia los cilios más pequeños, los canales se cierran y la célula se inhibe (hiperpolarizada).
- Torsión
- Un movimiento rotacional del ojo alrededor de la línea de visión que consiste en un sentido horario o antihorario.
- Ángulo de vergencia
- El ángulo entre la línea de visión para los dos ojos. Los ángulos de vergencia bajos indican objetos de visión lejana, mientras que los ángulos grandes indican la visualización de objetos cercanos.
- Compensación vestibular
- Después de una lesión en un lado de los receptores vestibulocleares o del nervio vestibulococlear, las neuronas de los núcleos vestibulares centrales recuperan gradualmente gran parte de su función a través de mecanismos de plasticidad. Sin embargo, la recuperación nunca es completa y los entornos de movimiento extremo pueden provocar mareos, náuseas, problemas de equilibrio y memoria espacial.
- Eferentes vestibulares
- Fibras nerviosas originadas en un núcleo en el tronco encefálico que se proyectan desde el cerebro para inervar las células ciliadas del receptor vestibular y las terminales nerviosas aferentes. Los eferentes tienen un papel modulador en sus dianas, lo cual no se entiende bien.
- Sistema vestibular
- Consiste en un conjunto de receptores de detección de movimiento y gravedad en el oído interno, un conjunto de núcleos primarios en el tronco encefálico y una red de vías que transportan señales de movimiento y gravedad a muchas regiones del cerebro.
- Nervio vestibulococlear
- El VIIIo nervio craneal que porta fibras inervando los receptores vestibulares y la cóclea.
- Reflejo vestibuloocular
- Movimientos oculares producidos por el tronco encefálico vestibular que son iguales en magnitud y opuestos en dirección al movimiento de la cabeza. El VOR funciona para mantener la estabilidad visual en un punto de interés y es casi perfecto para todos los movimientos naturales de la cabeza.
Referencias
- Angelaki, D. E., McHenry, M. Q., & Hess, B. J. (2000). Reflejos vestibulooculares traslacionales de primates. I. Dinámica de alta frecuencia y propiedades tridimensionales durante el movimiento lateral. J. Neurofisiol 83, 1637—1647.
- Angelaki, D. E., Shaikh, A. G., Green, A. M., & Dickman, J. D. (2004). Las neuronas computan modelos internos de las leyes físicas del movimiento. Naturaleza, 430, 560—564.
- Anniko, M., & Arnold, W. (1991). Localización del receptor de acetilcolina en células ciliadas cocleares y vestibulares adultas humanas. AC y Otolar, 111, 491—499.
- Beraneck, M., Hachemaoui, M., Idoux, E., Ris, L., Uno, A., Godaux, E.,. Vibert, N. (2003). Plasticidad a largo plazo de neuronas ipsilesionales del núcleo vestibular medial tras laberintiectomía unilateral. J Neurofisiol, 90, 184—203.
- Blanks, R.H.I., Curthoys, I. S., Bennett, M. L., & Markham, C. H. (1985). Relaciones planas de los canales semicirculares en monos rhesus y ardilla. Brain Res, 340, 315—324.
- Boyle, R., Carey, J. P., & Highstein, S.M. (1991). Correlatos morfológicos de dinámica de respuesta y estimulación eferente en aferentes de canal semicircular horizontal del pez sapo, Opsanus tau. J Neurofisiol, 66, 1504—1511.
- Brodal, A. Los núcleos vestibulares en el mono macaco. (1984). J. Comp. Neurol., 227, 252—266.
- Carleton, S. C., & Carpenter, M. B. (1984). Distribución de fibras vestibulares primarias en el tronco encefálico y cerebelo del mono. Brain Res, 294, 281—298.
- Chen, A., DeAngelis, G. C., & Angelaki, D. E. (2011). Comparación de la afinación espaciotemporal vestibular en la corteza vestibular parietoinsular de macaco, el área intraparietal ventral y el área temporal medial superior. J. Neurosci., 31, 3082—3094.
- Crane, B. T., & Demer, J. L. (1997). Estabilización de la mirada humana durante actividades naturales: efectos de traslación, rotación, aumento y distancia objetivo. J Neurofisiol, 78, 2129—2144.
- Dickman, J. D., & Angelaki, D. E. (2002). Patrones de convergencia vestibular en neuronas de núcleos vestibulares de primates alertas. J. Neurofisiol., 88, 3518—3533.
- Dickman, J. D., & Correia, M. J. (1989). Respuestas de fibras aferentes de canal semicircular horizontal de paloma. I. Estimulación mecánica y rotacional sinusoidal escalonada, trapezoidal y de baja frecuencia. J Neurofisiol, 62, 1090—1101.
- Dickman, J. D., & Fang, Q. (1996). Proyecciones centrales diferenciales de aferentes vestibulares en palomas. J Comp Neurol, 367, 110—121.
- Einstein, A. (1907) .Sobre el principio de relatividad y las conclusiones extraídas del mismo. Jahrb. Radioakt, 4, 411—462.
- Ezure, K., & Graf, K.W. (1984). Análisis cuantitativo de la organización espacial de los reflejos vestibulo-oculares en animales de ojos laterales y frontales. I. Orientación de canales semicirculares y músculos extraoculares. Neurociencia, 12, 85—94.
- Fernández, C., & Goldberg, J. M. (1976b). Fisiología de las neuronas periféricas que inervan órganos otolitos del mono ardilla. II. Selectividad direccional y relaciones fuerza-respuesta. J. Neurofisiol., 39, 985—995.
- Fernández, C., & Goldberg, J. M. (1971). Fisiología de las neuronas periféricas que inervan canales semicirculares del mono ardilla. II. Respuesta a la estimulación sinusoidal y dinámica del sistema vestibular periférico. J Neurofisiol, 34, 661—675.
- Fernández, C., & Goldberg, J.M. (1976a). Fisiología de las neuronas periféricas que inervan órgano otolito en el mono ardilla. I. Respuesta a inclinaciones estáticas y a la fuerza centrífuga de larga duración. J. Neurofisiol., 39, 970—984.
- Fetter, M., & Zee, D. S. (1988). Recuperación de laberintectomía unilateral en mono rhesus. J Neurofisiol, 59, 370—393.
- Rebaño, A. (1964). Estructura de la mácula utriculi con especial referencia a la interacción direccional de las respuestas sensoriales reveladas por la polarización morfológica. J Cell Biol, 22, 413—431.
- Fukushima, K., Sato, T., Fukushima, J. Shinmei, Y., y Kaneko C.R. (2000). Actividad de neuronas relacionadas con la búsqueda suave en las cortes perarcuadas de mono durante la persecución y rotación pasiva de todo el cuerpo. J. Neurophys., 83, 563—587.
- Grusser, O., Pausa, M., & Schreiter, U. (1982). Respuestas neuronales en la corteza vestibular parieto-insular de monos java alertas (Macaca fascicularis). En A. Roucoux y M. Crommelinck (Eds.), Aspectos fisiológicos y patológicos de los movimientos oculares. El Haque: W. Junk Publishers.
- Gu, Y., DeAngelis, G. C., & Angelaki, D. E. (2007). Un vínculo funcional entre el área mstd y la percepción de rumbo basado en señales vestibulares. Nat Neurosci, 10, 1038—1047.
- Gu, Y., Watkins, P. V., Angelaki, D. E., & DeAngelis, G. C. (2006). Contribuciones visuales y no visuales a la selectividad de rumbo tridimensional en el área temporal media superior. J Neurosci, 26, 73—85.
- Guldin, W. O., Akbarian, S., & Grusser, O. J. (1992). Conexiones cortico-corticales y citoarquitectónica de la corteza vestibular de primates: Un estudio en monos ardilla (Saimiri sciureus). J. Comp. Neurol., 326, 375—401.
- Haffting, T., Fyhn, M., Molden, S., Moser, M.-B., Moser, E. I. (2005). Microestructura de un mapa espacial en la corteza entorinal. Naturaleza, 436, 801—806.
- Iurato, S. (1967). Estructura submicroscópica del oído interno. Londres, Inglaterra: Pergamon Press.
- Kasahara, M., & Uchino, Y. (1974). Entradas de canal semicircular bilateral a neuronas en núcleos vestibulares de gatos. Exp Brain Res, 20, 285—296.
- Keshner, E. A., & Peterson, B. W. (1995). Mecanismos que controlan la estabilización de cabeza humana. I. Dinámica cabeza-cuello durante rotaciones aleatorias en el plano horizontal. J Neurofisiol, 73, 2293—2301.
- Lindeman, H. H. (1969). Estudios sobre la morfología de las regiones sensoriales del aparato vestibular con 45 figuras. Ergeb Anat Entwicklungsgesch, 42, 1—113.
- Lisberger, S. G., Pavelko, T. A., & Broussard, D. M. (1994). Bases neuronales para el aprendizaje motor en el reflejo vestibuloocular de primates. I. Cambios en las respuestas de las neuronas del tronco encefálico. J. Neurofisiol, 72, 928—973.
- Marlinski, V., & McCrea, R. A. (2008). Actividad de las neuronas del tálamo ventroposterior durante la rotación y traslación en el plano horizontal en el mono ardilla alerta. J Neurofisiol, 99, 2533—2545.
- McNaughton, B. L., Battaglia, F. P., Jensen, O., Moser, E. I., & Moser, M.-.B. (2006). La integración de caminos y la base neuronal del “mapa cognitivo”. Nature Rev., 7, 663—678.
- Meng, H., May, P. J., Dickman, J. D., & Angelaki, D. E. (2007). Señales vestibulares en tálamo de primates: Propiedades y orígenes. J Neurosci, 27, 13590—13602.
- Newlands, S. D., & Perachio, A. A. (1990). Compensación de la actividad relacionada con el canal horizontal en el núcleo vestibular medial tras la ablación laberíntica unilateral en el jerbo descerebrado. i. neuronas tipo i. Exp Brain Res, 82, 359—372.
- Newlands, S. D., Hesse, S. V., Haque, A., & Angelaki, D. E. (2001). La mirada horizontal descontrolada de la cabeza se desplaza tras laberintiectomía unilateral en el mono rhesus. Exp Brain Res, 140, 25—33.
- O'Keefe, J. (1976). Colocar unidades en el hipocampo de la rata que se mueve libremente. Exp Neurol, 51, 78—109.
- Paige, G. D., & Tomko, D. L. (1991). Respuestas del movimiento ocular al movimiento lineal de la cabeza en el mono ardilla. I. Características básicas. J Neurofisiol, 65, 1170—1182.
- Peterson, B. W., Goldberg, J., Bilotto, G., & Fuller, J. H. (1985). Reflejo cervicocólico: Sus propiedades dinámicas e interacción con reflejos vestibulares. J Neurofisiol, 54, 90—109.
- Precht, W., & Shimazu, H. (1965). Conexiones funcionales de neuronas vestibulares tónicas y cinéticas con aferentes vestibulares primarios. J Neurofisiol, 28, 1014—1028.
- Sadeghi, S. G., Chacron, M. J., Taylor, M. C., & Cullen, K. E. (2007). Variabilidad neural, umbrales de detección y transmisión de información en el sistema vestibular. J. Neurosci., 27, 771—781.
- Schwarz, U., Busettini, C., & Miles, F. A. (1989). Las respuestas oculares al movimiento lineal son inversamente proporcionales a la distancia de visión. Ciencia, 245, 1394-1396.
- Shimazu, H., & Precht, W. (1966). Inhibición de neuronas vestibulares centrales del laberinto contralateral y su vía mediadora. J Neurofisiol, 29, 467—492.
- Shinoda, Y., Sugiuchi, Y., Futami, T., Ando, N., & Kawasaki, T. (1994). Patrones de entrada y vías desde los seis canales semicirculares hasta las motoneuronas de los músculos del cuello. I. El grupo de músculos multifidus. J. Neurofisiol., 72, 2691—2802.
- Shotwell, S. L, Jacobs, R., & Hudspeth, A. J. (1981). Sensibilidad direccional de las células ciliadas de vertebrados individuales a la deflexión controlada de sus haces capilares. ANYAS, 374, 1—10.
- Stackman, R. W., Clark, A. S., & Taube, J. (2002). Las representaciones espaciales hipocampales requieren entrada vestibular. Hipocampo, 12, 291—303.
- Taube, J .S. (1995). Células en dirección de cabeza registradas en los núcleos talámicos anteriores de ratas que se mueven libremente. J. Neurosci., 15, 70—86.
- Van Egmond, A. A. J., Groen, J. J., & Jongkess, L. B. W. (1949). La mecánica de los canales semicirculares. J Physiol Lond, 110, 1—17.
- Wersäll, J. (1956). Estudios sobre la estructura e inervación del epitelio sensorial de las cristae ampullares en el cobaya; una investigación microscópica de luz y electrónica. Acta Otorrinolaringol Supp, 126, 1—85.
- Wilson, V. J., Boyle, R., Fukushima, K., Rose, P. K., Shinoda, Y., Sugiuchi, Y., & Uchino, Y. (1995). El reflejo vestibulocólico. J Vestib Res, 5, 147—170.
- Yates, B. J., Miller A. D., & Lucot, J. B. (1998). Bases fisiológicas y farmacología del mareo por movimiento: Una actualización. Brain Res Bull, 47, 395—406.
- Yu, X. J., Dickman, J. D., Angelaki, D. E. (2012). Umbrales de detección de aferentes de otolito de macaco. J Neurosci., 32, 8306—8316.
- Zhou, W., Tang, B.F., Newlands, S.D., King, W.M. (2006). Respuestas de neuronas solo vestibulares de mono a la traslación y rotación angular. J Neurofisiol 96, 2915-2930.