
5.3: Visión
- Page ID
- 149298

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
Anatomía del Sistema Visual
El ojo es el principal órgano sensorial involucrado en la visión (Figura 5.11). Las ondas de luz se transmiten a través de la córnea y entran al ojo a través de la pupila. La córnea es la cubierta transparente sobre el ojo. Sirve como barrera entre el ojo interno y el mundo exterior, y se involucra en enfocar las ondas de luz que ingresan al ojo. La pupila es la pequeña abertura en el ojo por la que pasa la luz, y el tamaño de la pupila puede cambiar en función de los niveles de luz así como de la excitación emocional. Cuando los niveles de luz son bajos, la pupila se dilatará, o se expandirá, para permitir que entre más luz en el ojo. Cuando los niveles de luz son altos, la pupila se contraerá, o se volverá más pequeña, para reducir la cantidad de luz que ingresa al ojo. El tamaño de la pupila está controlado por músculos que están conectados al iris, que es la porción coloreada del ojo.
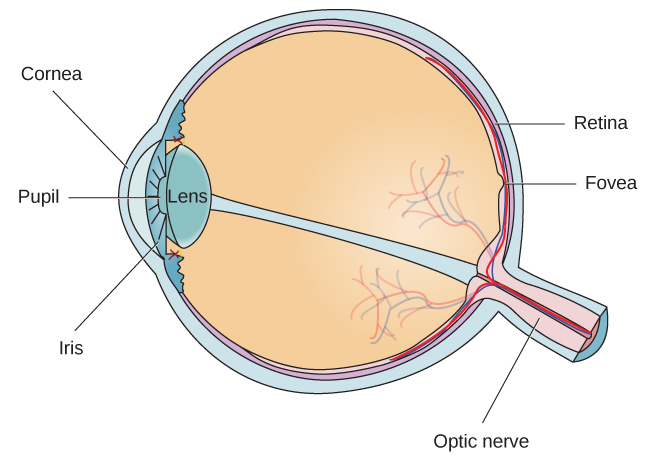
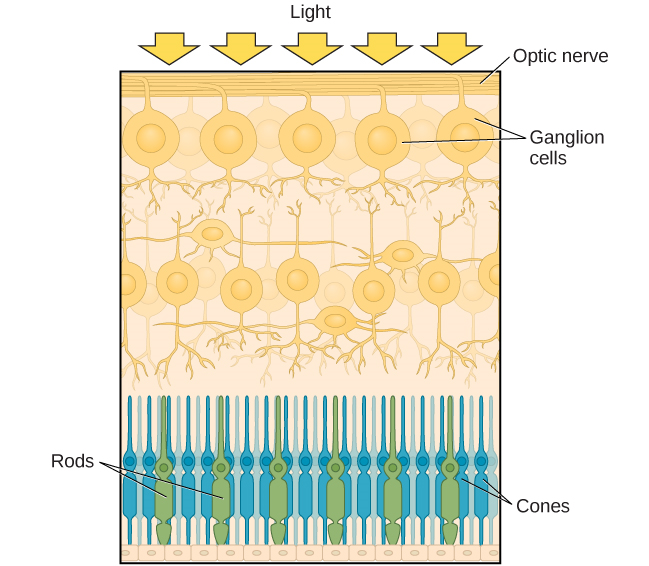
Después de pasar por la pupila, la luz cruza la lente, una estructura curva y transparente que sirve para proporcionar un enfoque adicional. La lente está unida a músculos que pueden cambiar su forma para ayudar a enfocar la luz que se refleja desde objetos cercanos o lejanos. En un individuo de visión normal, el cristalino enfocará las imágenes perfectamente en una pequeña muesca en la parte posterior del ojo conocida como la fóvea, que es parte de la retina, el revestimiento sensible a la luz del ojo. La fóvea contiene células fotorreceptoras especializadas densamente empaquetadas (Figura 5.12). Estas células fotorreceptoras, conocidas como conos, son células detectoras de luz. Los conos son tipos especializados de fotorreceptores que funcionan mejor en condiciones de luz brillante. Los conos son muy sensibles a los detalles agudos y proporcionan una tremenda resolución espacial. También están directamente involucrados en nuestra capacidad de percibir el color.
Mientras que los conos se concentran en la fóvea, donde las imágenes tienden a enfocarse, los bastones, otro tipo de fotorreceptores, se localizan a lo largo del resto de la retina. Las varillas son fotorreceptores especializados que funcionan bien en condiciones de poca luz, y si bien carecen de la resolución espacial y la función de color de los conos, están involucradas en nuestra visión en ambientes poco iluminados así como en nuestra percepción del movimiento en la periferia de nuestro campo visual.
Todos hemos experimentado las diferentes sensibilidades de los bastones y conos al hacer la transición de un ambiente con mucha luz a un ambiente con poca luz. Imagina ir a ver una película de gran éxito en un claro día de verano. Al caminar desde el vestíbulo brillantemente iluminado hacia el teatro oscuro, nota que inmediatamente tiene dificultades para ver mucho de cualquier cosa. Después de unos minutos, comienzas a ajustarte a la oscuridad y podrás ver el interior del teatro. En el ambiente brillante, tu visión estuvo dominada principalmente por la actividad de cono. A medida que te mueves al ambiente oscuro, la actividad de la vara domina, pero hay un retraso en la transición entre las fases. Si tus bastones no transforman la luz en impulsos nerviosos tan fácil y eficientemente como deberían, tendrás dificultades para ver con poca luz, condición conocida como ceguera nocturna.
Los bastones y conos están conectados (a través de varias interneuronas) a células ganglionares retinianas. Los axones de las células ganglionares de la retina convergen y salen por la parte posterior del ojo para formar el nervio óptico. El nervio óptico transporta información visual desde la retina hasta el cerebro. Hay un punto en el campo visual llamado punto ciego: Incluso cuando la luz de un objeto pequeño se enfoca en el punto ciego, no lo vemos. No somos conscientes de nuestros puntos ciegos por dos razones: Primero, cada ojo obtiene una visión ligeramente diferente del campo visual; por lo tanto, los puntos ciegos no se superponen. Segundo, nuestro sistema visual rellena el punto ciego para que aunque no podamos responder a la información visual que se produce en esa parte del campo visual, tampoco somos conscientes de que falta información.
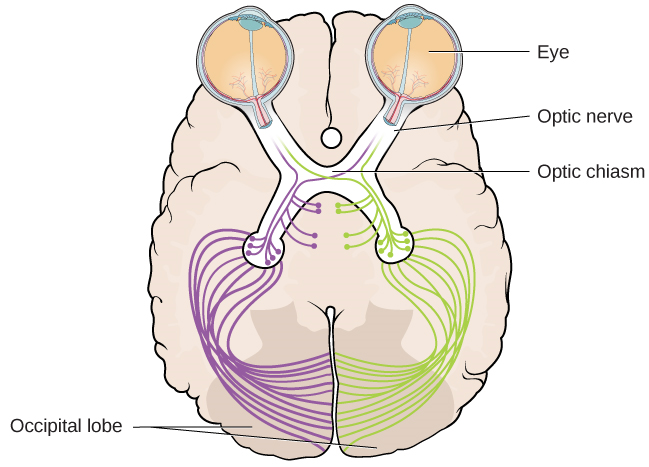
El nervio óptico de cada ojo se funde justo debajo del cerebro en un punto llamado quiasma óptico. Como muestra la Figura 5.13, el quiasma óptico es una estructura en forma de X que se asienta justo debajo de la corteza cerebral en la parte frontal del cerebro. En el punto del quiasma óptico, la información del campo visual derecho (que proviene de ambos ojos) se envía al lado izquierdo del cerebro, y la información del campo visual izquierdo se envía al lado derecho del cerebro.
La ética de la investigación con animales
David Hubel y Torsten Wiesel fueron galardonados con el Premio Nobel de Medicina en 1981 por sus investigaciones sobre el sistema visual. Colaboraron durante más de veinte años e hicieron importantes descubrimientos sobre la neurología de la percepción visual (Hubel & Wiesel, 1959, 1962, 1963, 1970; Wiesel & Hubel, 1963). Estudiaron animales, en su mayoría gatos y monos. A pesar de que utilizaron varias técnicas, realizaron considerables registros de una sola unidad, durante los cuales se insertaron pequeños electrodos en el cerebro del animal para determinar cuándo se activó una sola célula. Entre sus muchos descubrimientos, encontraron que células cerebrales específicas responden a líneas con orientaciones específicas (llamadas dominancia ocular), y mapearon la forma en que esas células están dispuestas en áreas de la corteza visual conocidas como columnas e hipercolumnas.
En algunas de sus investigaciones, suturaron un ojo de gatitos recién nacidos cerrados y siguieron el desarrollo de la visión de los gatitos. Descubrieron que había un periodo crítico de desarrollo para la visión. Si a los gatitos se les privaba de entrada de un ojo, otras áreas de su corteza visual llenaban el área que normalmente usaba el ojo que estaba cosido cerrado. En otras palabras, las conexiones neuronales que existen al nacer pueden perderse si se les priva de la entrada sensorial.
¿Qué opinas sobre coser un ojo de gatito cerrado para investigación? A muchos defensores de los animales, esto parecería brutal, abusivo y poco ético. ¿Y si pudieras hacer investigaciones que ayuden a garantizar que los bebés y niños nacidos con ciertas afecciones puedan desarrollar una visión normal en lugar de quedar ciegos? ¿Quieres que se haga esa investigación? ¿Realizarías esa investigación, aunque signifique causar algún daño a los gatos? ¿Pensarías de la misma manera si fueras el padre de un niño así? ¿Y si trabajaste en el refugio de animales?
Como prácticamente cualquier otra nación industrializada, Estados Unidos permite la experimentación médica en animales, con pocas limitaciones (asumiendo suficiente justificación científica). El objetivo de cualquier ley que exista no es prohibir tales pruebas sino limitar el sufrimiento innecesario de los animales estableciendo normas para el trato humano y alojamiento de los animales en los laboratorios.
Según explicó Stephen Latham, director del Centro Interdisciplinario de Bioética de Yale (2012), los posibles enfoques legales y regulatorios de las pruebas con animales varían en un continuo desde una fuerte regulación gubernamental y monitoreo de toda experimentación en un extremo, hasta un enfoque autorregulado que depende de la ética de los investigadores en el otro extremo. El Reino Unido tiene el esquema regulatorio más significativo, mientras que Japón utiliza el enfoque de autorregulación. El enfoque de Estados Unidos está en algún lugar en el medio, el resultado de una mezcla gradual de los dos enfoques.
No cabe duda de que la investigación médica es una práctica valiosa e importante. La cuestión es si el uso de animales es una práctica necesaria o incluso una mejor práctica para producir los resultados más confiables. Las alternativas incluyen el uso de bases de datos paciente-fármaco, ensayos virtuales de drogas, modelos y simulaciones por computadora, y técnicas de imagen no invasivas como imágenes por resonancia magnética y tomografía computarizada (“Animales en ciencia/Alternativas”, n.d.). Otras técnicas, como la microdosificación, utilizan a los humanos no como animales de prueba sino como un medio para mejorar la precisión y confiabilidad de los resultados de las pruebas. Los métodos in vitro basados en cultivos de células y tejidos humanos, células madre y métodos de prueba genética también están cada vez más disponibles.
Hoy en día, a nivel local, cualquier instalación que utilice animales y reciba financiamiento federal debe contar con un Comité Institucional de Cuidado y Uso de Animales (IACUC) que asegure que se sigan los lineamientos de los NIH. El IACUC debe incluir investigadores, administradores, un veterinario, y al menos una persona sin vinculación con la institución: es decir, un ciudadano preocupado. Este comité también realiza inspecciones de laboratorios y protocolos.
Percepción de Color y Profundidad
No vemos el mundo en blanco y negro; tampoco lo vemos como bidimensional (2-D) o plano (solo altura y ancho, sin profundidad). Veamos cómo funciona la visión del color y cómo percibimos tres dimensiones (alto, ancho y profundidad).
Visión del color
Los individuos con visión normal tienen tres tipos diferentes de conos que median la visión del color. Cada uno de estos tipos de cono es máximamente sensible a una longitud de onda de luz ligeramente diferente. De acuerdo con la teoría tricromática de la visión del color, mostrada en la Figura 5.14, todos los colores del espectro se pueden producir combinando rojo, verde y azul. Los tres tipos de conos son cada uno receptivos a uno de los colores.
El daltonismo: una historia personal
Hace varios años, me vestí para ir a una función pública y entré a la cocina donde estaba sentada mi hija de 7 años. Ella me miró, y con su voz más severa, dijo: “No puedes usar eso”. Yo le pregunté: “¿Por qué no?” y ella me informó que los colores de mi ropa no coincidían. Ella se había quejado frecuentemente de que yo era mala para combinar mis camisas, pantalones y corbatas, pero esta vez, ella sonó especialmente alarmada. Como padre soltero sin nadie más a quien preguntar en casa, nos llevé a la tienda de conveniencia más cercana y le pregunté al empleado de la tienda si mi ropa coincidía. Ella dijo que mis pantalones eran de color verde brillante, mi camisa era naranja rojiza y mi corbata era marrón. Ella me miró con interrogación y dijo: “De ninguna manera tu ropa coincide”. En los siguientes días, comencé a preguntar a mis compañeros de trabajo y amigos si mi ropa coincidía. Después de varios días de que me dijeron que mis compañeros de trabajo simplemente pensaban que tenía “un estilo realmente único”, concerté una cita con un oftalmólogo y me hicieron la prueba (Figura 5.15). Fue entonces cuando me enteré de que era daltónico. No puedo diferenciar entre la mayoría de los verdes, marrones y rojos. Afortunadamente, aparte de estar mal vestido sin saberlo, mi daltonismo rara vez daña mi día a día.
Algunas formas de deficiencia de color son raras. Ver en escala de grises (solo tonos de blanco y negro) es extremadamente raro, y las personas que lo hacen solo tienen varillas, lo que significa que tienen una agudeza visual muy baja y no pueden ver muy bien. La anomalía hereditaria ligada al X más común es el daltonismo rojo-verde (Birch, 2012). Aproximadamente el 8% de los hombres con decencia caucásica europea, el 5% de los asiáticos, el 4% de los hombres africanos y menos del 2% de los machos indígenas americanos, los machos australianos y los machos polinesios tienen deficiencia de color rojo-verde (Birch, 2012). Comparativamente, solo alrededor de 0.4% en las hembras de ascendencia caucásica europea tienen deficiencia de color rojo-verde (Birch, 2012).

Percepción de profundidad
Nuestra capacidad para percibir relaciones espaciales en el espacio tridimensional (3-D) se conoce como percepción de profundidad. Con la percepción de profundidad, podemos describir las cosas como estar al frente, detrás, arriba, abajo, o al lado de otras cosas.
Nuestro mundo es tridimensional, por lo que tiene sentido que nuestra representación mental del mundo tenga propiedades tridimensionales. Utilizamos una variedad de señales en una escena visual para establecer nuestro sentido de profundidad. Algunas de estas son señales binoculares, lo que significa que dependen del uso de ambos ojos. Un ejemplo de una señal de profundidad binocular es la disparidad binocular, la visión ligeramente diferente del mundo que recibe cada uno de nuestros ojos. Para experimentar esta visión ligeramente diferente, haz este sencillo ejercicio: extiende tu brazo completamente y extiende uno de tus dedos y concéntrate en ese dedo. Ahora, cierra el ojo izquierdo sin mover la cabeza, luego abre el ojo izquierdo y cierra el ojo derecho sin mover la cabeza. Notarás que tu dedo parece cambiar a medida que alternas entre los dos ojos debido a la vista ligeramente diferente que cada ojo tiene de tu dedo.
Una película 3-D funciona según el mismo principio: las gafas especiales que usas permiten que las dos imágenes ligeramente diferentes proyectadas en la pantalla sean vistas por separado por tu ojo izquierdo y derecho. A medida que tu cerebro procesa estas imágenes, tienes la ilusión de que el animal saltando o la persona que corre viene directo hacia ti.
Aunque confiamos en señales binoculares para experimentar profundidad en nuestro mundo 3D, también podemos percibir profundidad en matrices 2-D. Piensa en todas las pinturas y fotografías que has visto. Generalmente, captas profundidad en estas imágenes a pesar de que el estímulo visual es 2-D. Cuando hacemos esto, estamos confiando en una serie de señales monoculares, o señales que requieren solo un ojo. Si crees que no puedes ver la profundidad con un ojo, ten en cuenta que no te encuentras con las cosas cuando usas solo un ojo al caminar y, de hecho, tenemos más señales monoculares que binoculares.
Un ejemplo de una señal monocular sería lo que se conoce como perspectiva lineal. La perspectiva lineal se refiere al hecho de que percibimos profundidad cuando vemos dos líneas paralelas que parecen converger en una imagen (Figura 5.17). Algunas otras señales de profundidad monocular son la interposición, la superposición parcial de objetos y el tamaño relativo y cercanía de las imágenes al horizonte.

Bruce Bridgeman nació con un caso extremo de ojo vago que resultó en que fuera estereocigado, o incapaz de responder a señales binoculares de profundidad. Confió en gran medida en las señales de profundidad monocular, pero nunca tuvo una verdadera apreciación de la naturaleza 3-D del mundo que lo rodeaba. Todo esto cambió una noche de 2012 mientras Bruce estaba viendo una película con su esposa.
La película que la pareja iba a ver se rodó en 3-D, y a pesar de que pensó que era una pérdida de dinero, Bruce pagó las gafas 3-D cuando compró su boleto. Tan pronto como comenzó la película, Bruce se puso las gafas y experimentó algo completamente nuevo. Por primera vez en su vida apreció la verdadera profundidad del mundo que le rodea. Sorprendentemente, su capacidad para percibir la profundidad persistió fuera del cine.
Hay células en el sistema nervioso que responden a señales de profundidad binocular. Normalmente, estas células requieren activación durante el desarrollo temprano para persistir, por lo que los expertos familiarizados con el caso de Bruce (y otros como el suyo) asumen que en algún momento de su desarrollo, Bruce debió haber experimentado al menos un momento fugaz de visión binocular. Fue suficiente para asegurar la supervivencia de las células en el sistema visual sintonizado a las señales binoculares. El misterio ahora es por qué Bruce tardó casi 70 años en activar estas células (Peck, 2012).