7.1: Una visión general de las vías metabólicas - Catabolismo
- Page ID
- 79229
Las células biológicas tienen una tarea desalentadora. Deben llevar a cabo 1000s de diferentes reacciones químicas requeridas para llevar a cabo la función celular. Estas reacciones pueden incluir objetivos opuestos como la producción de energía y almacenamiento de energía, la degradación y síntesis de macromoléculas, y la descomposición y síntesis de moléculas pequeñas. Todas estas reacciones son catalizadas por proteínas y enzimas de ARN cuyas actividades deben regularse, nuevamente a través de reacciones químicas, para evitar un escenario inútil y de pérdida de energía de tener vías opuestas funcionando simultáneamente en una célula.
El metabolismo se puede dividir en dos partes principales, el catabolismo, la degradación de las moléculas, generalmente para producir energía o moléculas pequeñas útiles para la función celular, y el anabolismo, la síntesis de biomoléculas más grandes a partir de pequeños precursores.
CATBILIA: Las reacciones catabólicas implican la descomposición de carbohidratos, lípidos, proteínas y ácidos nucleicos para producir moléculas más pequeñas y energía biológica en forma de calor o pequeñas moléculas termodinámicamente reactivas como el ATP cuya degradación adicional puede impulsar procesos endergónicos como la biosíntesis. Todo nuestro mundo depende de la oxidación de hidrocarburos orgánicos a agua y dióxido de carbono para producir energía (a expensas de liberar un potente gas de efecto invernadero,\(\ce{CO2}\)). En el mundo biológico, las moléculas reducidas como los ácidos grasos y las moléculas parcialmente oxidadas como los polímeros de glucosa (glucógeno, almidón), así como los azúcares simples, pueden oxidarse parcial o completamente para finalmente producir\(\ce{CO2}\) también. La energía liberada por las reacciones oxidativas se utiliza para producir moléculas como ATP así como calor. Las vías oxidativas incluyen la glucólisis, el ciclo del ácido tricarboxílico (también conocido como el ciclo de Kreb) y la fosforilación oxidativa mitocondrial/transporte de electrones. Para oxidar completamente el carbono en la glucosa y los ácidos grasos a dióxido de carbono requiere dividir los enlaces C-C y la disponibilidad de series de agentes oxidantes que puedan realizar reacciones de oxidación controladas y escalonadas, análogas a la oxidación secuencial de metano, CH4 a metanol (CH3OH), formaldehído (CH2O) y carbono dixóxido.
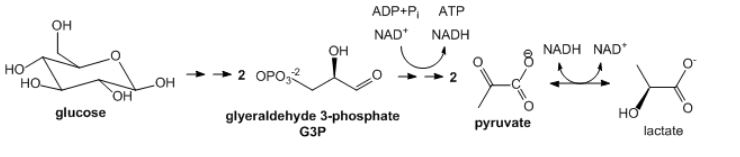
- Glicólisis: Esta vía metabólica más primitiva se encuentra quizás en todos los organismos. En la glucólisis, la glucosa (\(\ce{C6H12O6}\)), una molécula 6C, se divide (o se lisa) en dos moléculas de carbono 3C, gliceraldehído-3-fosfato, que luego se oxidan parcialmente en condiciones anaerobias (sin\(\ce{O2}\)) para formar dos moléculas de piruvato (\(\ce{CH3COCO2^{-}}\)). En lugar del agente oxidante muy fuerte\(\ce{O2}\), uno más débil, se usa NAD+, el cual se reduce en el proceso para formar NADH. Dado que ninguno de los átomos de carbono se oxida al estado de\(\ce{CO2}\), se libera poca energía en comparación con la oxidación completa a\(\ce{CO2}\). Esta vía llega a un alto chillido si todo el NAD+ celular se convierte en NADH ya que el NAD+ no se repone por el simple acto de respirar como es el caso\(\ce{O2}\) en la oxidación aeróbica. Para evitar que el agotamiento de NAD+ inhiba el ciclo y permitir que el ciclo continúe en condiciones anaerobias, el exceso de NADH se reconvierte a NAD+ cuando el otro producto de la glucólisis, el piruvato se convierte en lactato por la enzima lactato deshidrogenasa. La glucólisis ocurre en el citoplasma de la célula.
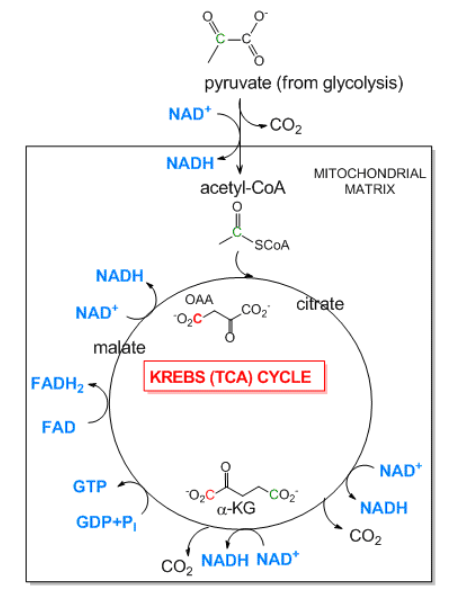
- Ciclo del ácido tricarboxílico (Kreb's): El ciclo de TCA es una vía aeróbica que tiene lugar en un orgánulo intracelular llamado mitocondrias. Toma piruvato, el producto incompletamente oxidado de la glucólisis, y termina el trabajo de oxidar los átomos 3C hasta el final\(\ce{CO2}\). Primero el piruvato se mueve hacia las mitocondrias donde se oxida a la molécula 2C acetilCoA con la liberación de una\(\ce{CO2}\) por la enzima piruvato deshidrogenasa. El acetil-CoA luego ingresa al ciclo de TCA donde\(\ce{CO2}\) se liberan dos más. Al igual que en la glucólisis, los enlaces C-C se escinden y el C es oxidado por NAD+ y otro agente oxidante relacionado, FAD. Lo que es muy diferente de esta vía es que en lugar de ser una serie de reacciones lineales y secuenciales con un reactivo (glucosa) y un producto (dos pryuvatos), es una vía cíclica. Esto tiene consecuencias significativas ya que si alguno de los reactivos dentro de las vías se agota, toda la ruta cíclica puede ralentizarse y detenerse. Para ver cómo sucede esto considera la molécula oxaloacetato (OAA) que se condensa con acetil-CoA para formar citrato (ver diagrama a continuación). En esta reacción, se consume una OAA. Sin embargo, cuando el ciclo regresa, un malato se convierte en OAA por lo que no hay pérdida neta de OAA, a menos que se saque OAA del ciclo de TCA para otras reacciones, lo que sucede.
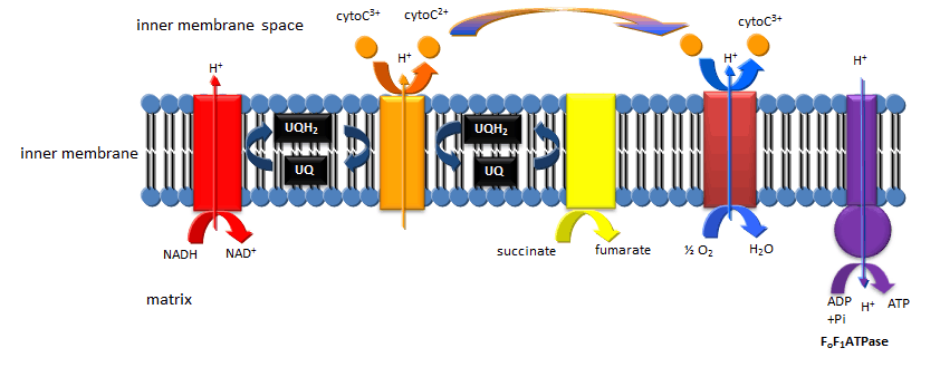
- Fosforilación Oxidativa Mitocondrial/Transporte de Electrones: El ciclo de TCA logra lo que la glucólisis no logró, es decir, la escisión de todos los enlaces C-C en la glucosa (en forma de piruvato y acetil-CoA, y la oxidación completa de todos los átomos de C a\(\ce{CO2}\). Sin embargo, quedan dos problemas. El conjunto de moléculas oxidantes, NAD+ y FAD se convierten a sus formas reducidas, NADH y FADH2. A menos que el NAD+ y el FAD se regeneren, como fue el caso en condiciones anaeróbicas cuando el piruvato se convierte en lacato, la vía volvería a detenerse en la molienda. Además, no se hace mucho ATP en el ciclo (en forma de una molécula relacionada GTP). Ambos problemas se resuelven ya que el NADH y el FADH2 resultantes formados son reoxidados por complejos enzimáticos de membrana mitocondrial que pasan electrones del NADH y FADH2 oxidado a agentes oxidantes cada vez más potentes hasta que son aceptados por el poderoso oxidante\(\ce{O2}\), que se convierte reducido a agua. La oxidación neta de NADH y FADH2 por dioxígeno es muy exergónica, y la energía liberada por el proceso impulsa la síntesis de ATP a partir de ADP y Pi por un complejo enzimático mitocondrial, la F0F1ATPasa.
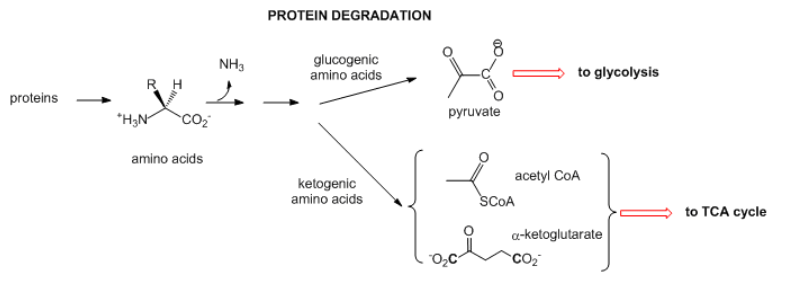
Vías Alimentadoras: Otras vías catabólicas producen productos que pueden ingresar a la glucólisis o al ciclo de TCA. A continuación se dan dos ejemplos.
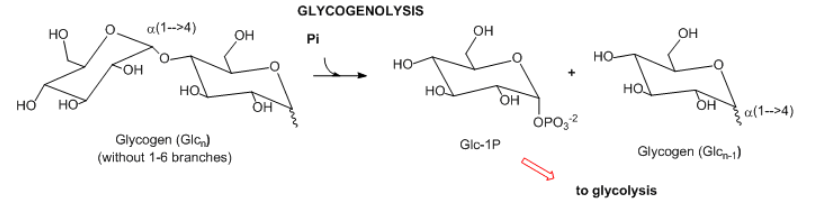
- Carbohidratos complejos: En los mamíferos, la principal molécula de almacenamiento de carbohidratos es el glucógeno, un polímero de glucosa unido a1-4 con ramas a1-6. Los enlaces acetales terminales en este polímero altamente ramificado se escinden secuencialmente en los extremos no a través de hidrólisis sino a través de fosforólisis para producir lotes de glucosa-1-fosfato que pueden ingresar a la glucólisis.
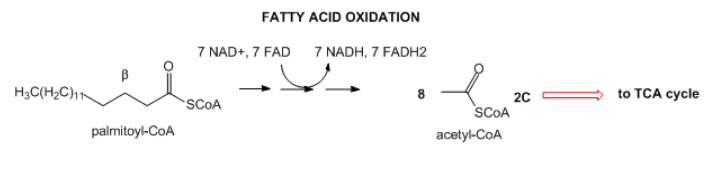
- Lípidos: Los lípidos se almacenan principalmente como triacilglicéridos en las células grasas (adipocitos). Cuando es necesario para obtener energía, los ácidos grasos se hidrolizan de la cadena principal de glicerol del triacilglicérido, y se envían a las células donde se descomponen en un proceso oxidativo para formar acetil-CoA con la producción concomitante de lotes de NADH y FADH2. Estos pueden entonces ingresar al sistema mitocondrial oxidativo de fosforilación/transporte de electrones, que produce, bajo condiciones aeróbicas, mucho ATP.
- Proteínas: Cuando las proteínas intracelulares se degradan, proceden de aminoácidos individuales. La amina N se pierde a medida que entra en el ciclo de la urea. El resto de algunas estructuras de aminoácidos se pueden convertir finalmente en acetil-CoA o cetoácidos (como alfa-cetoglutarato- A-kg) que son intermedios de TCA. Estos aminoácidos se llaman cetogénicos. Alternativamente, algunos aminoácidos, después de la desaminación, son codiciados para piruvato que puede entrar en el ciclo de TCA o en el hígado ser utilizado para sintetizar glucosa en un proceso anabólico. Estos aminoácidos se llaman glucógenos. Las reacciones químicas como estas pueden usarse para reponer intermedios en el ciclo de TCA que pueden agotarse a medida que se retiran para otras reacciones.