
7.2: Una visión general de las vías metabólicas - Anabolismo
- Page ID
- 79230
Anabolismo: Las reacciones anabólicas son aquellas que conducen a la síntesis de biomoléculas. En contraste con las reacciones catabólicas recién discutidas (glucólisis, ciclo de TCA y transporte de electrones/fosforilación oxidativa) que conducen a la degradación oxidativa de carbohidratos y ácidos grasos y liberación de energía, las reacciones anabólicas conducen a la síntesis de biomoléculas más complejas incluyendo biopolímeros ( glucógeno, proteínas, ácidos nucleicos) y lípidos complejos. Muchas reacciones biosintéticas, incluidas las de síntesis de ácidos grasos, son reductoras y, por lo tanto, requieren agentes reductores. La biosíntesis reductora y la formación de polímeros complejos requieren un aporte de energía, generalmente en forma de ATP cuya escisión exergónica se acopla a la biosíntesis endergónica.
Las células han desarrollado un mecanismo interesante para no tener reacciones de degradación oxidativa (que liberan energía) procedan al mismo tiempo y en la misma célula que la biosíntesis reductora (que requiere aporte de energía). Considera este escenario. Te sumerjas en una célula hepática y encuentras ácido palmítico, un ácido graso 16C. ¿De dónde vino? ¿Se acaba de sintetizar por la célula hepática o simplemente ingresó a la célula desde una ubicación distante como los adipocitos (células grasas). ¿Debería oxidarse, lo que debería suceder si existe una demanda de producción de energía por parte de la célula, o la célula hepática debería exportarla, tal vez a los adipocitos, lo que podría suceder si hay un exceso de moléculas de almacenamiento de energía? Las células han ideado muchas formas de distinguir estas necesidades opuestas. Una es mediante el uso de un conjunto ligeramente diferente de reactivos redox para reacciones anabólicas y catabólicas. Las reacciones de degradación oxidativa suelen utilizar el par redox NAD+/NADH (o FAD/FADH2) mientras que la biosíntesis reductora a menudo usa variantes fosforiladas de NAD+, NADP+/NADPH. Además, las células suelen llevar a cabo reacciones competitivas en diferentes compartimentos celulares. La oxidación de ácidos grasos de nuestra molécula de ejemplo (ácido palmítico) ocurre en la matriz mitocondrial, mientras que la síntesis reductiva de ácidos grasos ocurre en el citoplasma de la célula. Los ácidos grasos que ingresan a la célula destinados a la degradación oxidativa son transportados a las mitocondrias por el sistema de transporte de carnitina. Este sistema de transporte se inhibe en condiciones en las que se favorece la síntesis de ácidos grasos. Discutiremos la regulación de las vías metabólicas en una sección posterior. Uno de los principales métodos, como veremos, es activar o inhibir enzimas clave en las vías bajo un conjunto dado de condiciones celulares. La enzima clave en la síntesis de ácidos grasos, la acetil-CoA carboxilasa, se inhibe cuando las condiciones celulares requieren oxidación de ácidos grasos.
Los siguientes ejemplos dan breves descripciones de las vías anabólicas. Compárelas con las vías catabólicas de la sección anterior.
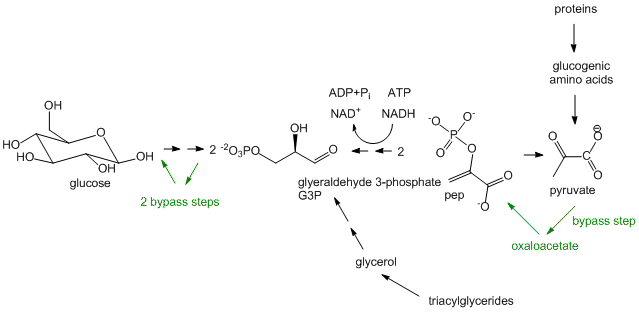
- Síntesis de glucosa, mejor conocida como Gluconeogénesis: En la glucólisis, la glucosa (C6H12O6), una molécula 6C, se convierte en dos moléculas 3C (piruvato) en un proceso oxidativo que requiere NAD+ y produce dos moléculas netas de ATP. En algunos órganos, sobre todo en el hígado, puede tener lugar la vía inversa. El hígado hace esto para proporcionar glucosa al cerebro cuando el cuerpo es deficiente en glucosa circulante, por ejemplo, bajo condiciones de ayuno y hambre. (El hígado en estas condiciones puede obtener su energía de la oxidación de los ácidos grasos). Las reacciones en la gluconeogénesis son las mismas reacciones en la glucólisis pero se ejecutan a la inversa, con la excepción de tres etapas glicolíticas que son esencialmente irreversibles. Estos tres pasos tienen enzimas de derivación en la vía de la gluconeogénesis. Aunque la síntesis de glucosa es una vía reductora, utiliza NADH en lugar de NADPH como redundante ya que la misma enzima utilizada en la glucólisis simplemente se ejecuta a la inversa. La gluconeogénesis, que también ocurre en la corteza del riñón, es más que una simple inversión de la glucólisis, sin embargo. Se puede considerar como la síntesis neta de glucosa a partir de precursores no carbohidratos. El piruvato, como se ve en la sección sobre catabolismo, se puede formar a partir de la degradación de proteínas a aminoácidos glucógenos que pueden convertirse en piruvato. También se puede formar a partir de triacilglicéridos a partir de la molécula 3C glicerol formada y liberada de los adipocitos después de la hidrólisis de tres ácidos grasos a partir de triacilglicéridos. Sin embargo, en humanos, la glucosa no se puede hacer de manera neta a partir de ácidos grasos. Los ácidos grasos se pueden convertir en acetil-CoA por oxidación de ácidos grasos. El acetil-CoA resultante no puede formar piruvato ya que la enzima que cataliza la formación de acetil-CoA a partir de piruvato, piruvato deshidrogenasa, es irreversible y no se conoce ninguna reacción de bypass. El acetil-CoA puede ingresar al ciclo de TCA pero como la ruta es cíclica y avanza en una dirección, no se puede formar en forma neta oxaloacetato. Aunque el oxaloacetato se puede eliminar del ciclo de TCA y ser utilizado para formar fosfoenolpiuvato, un intermedio glicolítico, un acetil-CoA se condensa con un oxaloacetato para formar citrato que conduce de nuevo a un oxaloacetato. De ahí que los ácidos grasos no puedan convertirse en glucosa y otros azúcares de manera neta.
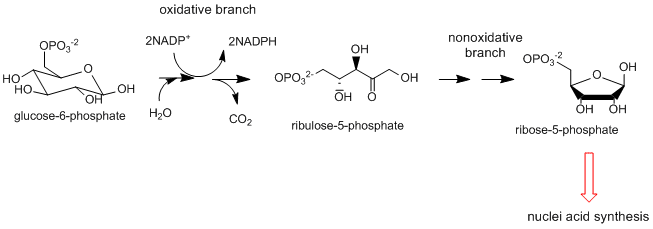
- Derivación de Pentosa Fosfato: Esta vía de dos partes no parece comenzar como una vía biosintética reductora ya que la primera parte es la conversión oxidativa de un intermedio glicolítico, glucosa-6-fosfato, a ribulosa-5-fosfato. La siguiente rama no oxidativa conduce a la formación de ribose-5-fosfato, un intermedio biosintético clave en la síntesis de ácidos nucleicos, así como ertirosa-4-fosfato utilizado para la biosíntesis de aminoácidos aromáticos. La rama oxidativa es importante en la biosíntesis reductora ya que es una fuente importante del reductor NADPH utilizado en reacciones biosintéticas.
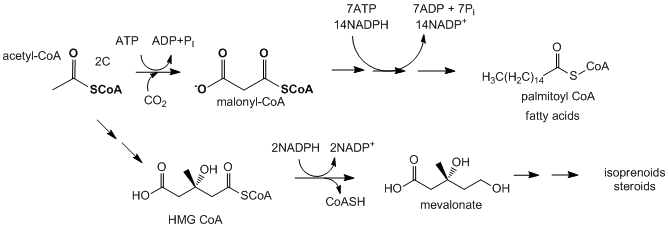
- Biosíntesis de ácidos grasos e isoprenoides/esterol: el acetil-CoA es la fuente de átomos de carbono para la síntesis de lípidos más complejos como ácidos grasos, isoprenoides y esteroles. Cuando las necesidades de energía en una célula no son altas, el citrato, el producto de condensación del oxaloacetato y acetil-CoA en el ciclo de TCA, se acumula en la matriz mitocondrial. Luego es transportado por el transportador de citrato (una proteína de membrana mitocondrial interna) al citoplasma, donde se escinde de nuevo a oxaloacetato y acetil-CoA por la enzima citoplásmica citrato liasa. El oxaloacetato se devuelve a las mitocondrias por conversión primero a malato (reacción de reducción usando NADH), que puede regresar a las mitocondrias a través del transportador de malato, o posterior conversión a piruato, usando la enzima málica citosólica, que usa NADP+ para oxidar malato a piruvato que luego entra en las mitocondrias. El acetil-CoA formado en el citoplasma se puede usar en biosíntesis reductora usando NADPH como reductor para formar ácidos grasos, isoprenoides y esteroles. El NADPH para la reducción proviene de la rama oxidativa de la vía de pentosa fosfato y de la reacción catalizada por la enzima málica. Las células hepáticas aún pueden ejecutar la vía glucolítica ya que la relación NADH/NAD+ es baja en el citoplasma mientras que la relación NADPH/NADP+ es alta.
Ahora es el momento de ver cómo los diversos caminos encajan entre sí para formar un conjunto integrado de vías.