
7.4: Complejo III
- Page ID
- 79371
El Complejo III acepta electrones tanto del Complejo I como del Complejo II. Los electrones llegan en forma de ubiquinol, UQH2, que entrega dos electrones y dos protones y se convierte de nuevo en ubiquinona, UQ. La ubiquinona luego se traslada de regreso al Complejo I o Complejo II para recolectar más electrones.
- El Complejo III es el destino de los electrones que llegan tanto del Complejo I como del Complejo II.
- El complejo III utiliza la energía liberada en transferencias de electrones descendentes para bombear más protones a través de la membrana mitocondrial interna.
- El gradiente de protones a través de la membrana se usa para impulsar la formación de ATP en el Complejo V.
- El Complejo III envía entonces sus electrones al Complejo IV.
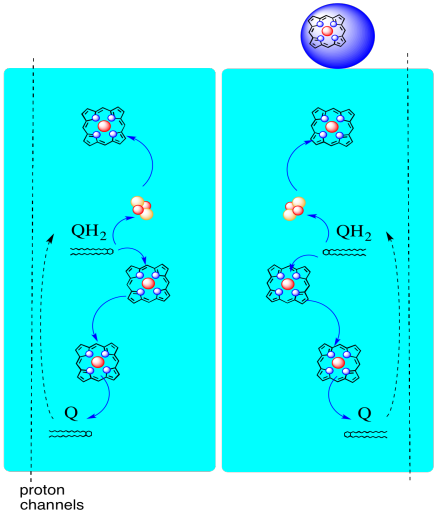
A continuación se muestra la trayectoria de los electrones a través del Complejo III. Obsérvese que el complejo es una estructura dimérica, con dos caminos equivalentes: uno que se muestra a la izquierda y el otro a la derecha. Los electrones se entregan desde UQH2, ubiquinol, en el medio de la imagen, viajando hacia arriba hasta el citocromo móvil c en la parte superior. Una segunda vía electrónica permite viajar hacia abajo en la imagen, hacia otra molécula de ubiquinona, que recicla los electrones en un bucle.
- El complejo III presenta una ruta electrónica inusual y divergente.
- Un electrón viaja hacia el citocromo c.
- El segundo electrón finalmente se recicla en otro ubiquinol.
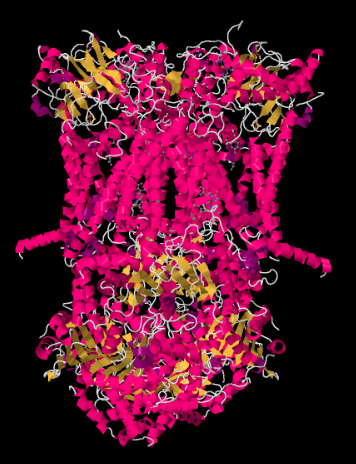
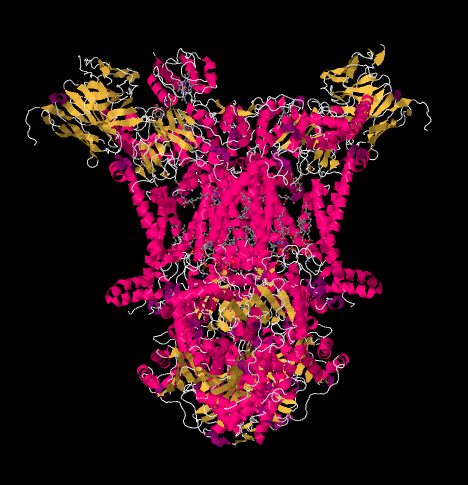
La estructura de rayos X del Complejo III se muestra en forma de caricatura a continuación. La membrana mitocondrial interna se cruzaría con la porción media superior del complejo. La mayoría del complejo se proyecta debajo, en la matriz (la parte inferior de la imagen, en esta orientación) aunque una cantidad significativa también sobresale en el espacio intermembrana.
El Complejo III continúa la cadena de transporte de electrones, enviando electrones a un mayor potencial de reducción (y menor energía libre) y hacia un encuentro con el oxígeno molecular en el Complejo IV. Los protones también se bombean desde la matriz mitocondrial, a través de la membrana mitocondrial interna y al espacio intermembranario. Al mismo tiempo, una segunda ruta de electrones da como resultado un reciclaje efectivo de la mitad de los electrones que entran en el complejo, aumentando el número de protones bombeados por electrón que llegan al Complejo III.
- Se ha sugerido que el bucle de reciclaje de electrones proporciona un mecanismo para extraer más protones de la matriz.
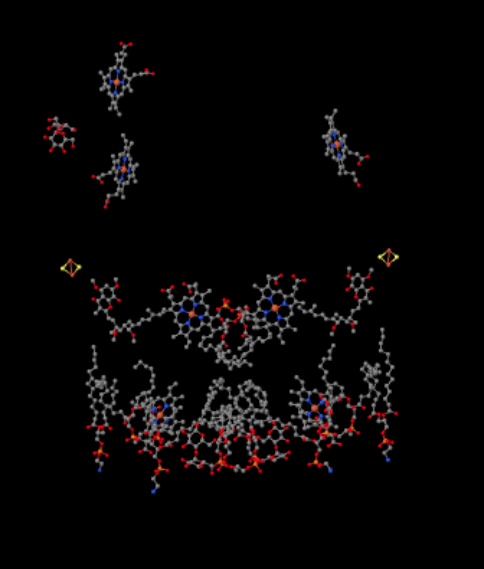
Podemos ver este arreglo inusual si ignoramos las proteínas para revelar los ligandos, a continuación. El complejo puede verse más claramente como un dímero; la mitad izquierda de la imagen es exactamente la misma que la derecha, aunque se refleja de manera opuesta. Los electrones ingresan al complejo a través del portador móvil, ubiquinol, visible en el lado derecho e izquierdo de la imagen aproximadamente a la mitad hacia abajo; se puede ver el anilloy una cadena colgando de él.
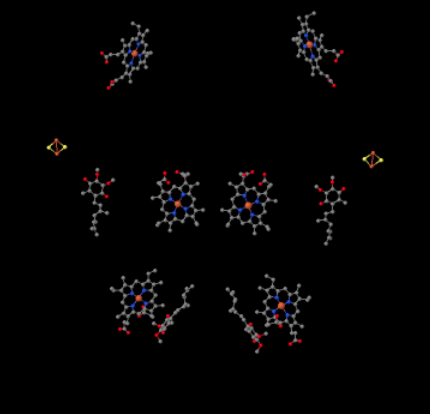
Cuando UQH2 se une al Complejo III, un electrón se envía a un grupo de FeS, mientras que el segundo se envía a un centro hemo de hierro, llamado hemo B L. El cúmulo de FeS es visible justo encima del ubiquinol y hasta el borde de la imagen. El hemo B L está hacia el centro de la imagen.
Aunque no es evidente en la imagen de rayos X que se muestra, este cúmulo de FeS es diferente a los que viste antes. Este se mantiene en su lugar con diferentes cadenas laterales de aminoácidos. No se puede ver eso en la imagen porque se ha dejado fuera toda la proteína, junto con aquellos aminoácidos que se están uniendo al cúmulo. Mientras que la mayoría de los racimos de FE se mantienen en su lugar exclusivamente por cisteínas, este racimo se mantiene en su lugar por dos cisteíneas en un lado pero por dos histidinas en el otro. Este peculiar cúmulo de FeS se llama racimo Rieske.
Los clústeres de Rieske son importantes porque son “clústeres de FE de alto potencial”. Eso significa que tienen potenciales de reducción excepcionalmente positivos en comparación con otros grupos de FE. El potencial de reducción de cualquier ligando en una proteína es altamente dependiente de su entorno, por lo que existe una amplia gama de valores en la mayoría de los casos. Eso es cierto para los grupos de hierro y azufre; los normales tienen potenciales de reducción que van de -1.0 V a aproximadamente +0.05 V. Los clústeres de Rieske tienen potenciales que varían de aproximadamente 0V a aproximadamente 0.4 V.
Desde el cúmulo de FeS, el electrón se envía a otro centro hemo de hierro en una unidad unida a membrana llamada citocromo c1. El citocromo c1 se puede ver en la parte superior de la imagen. Debido a que esta imagen está orientada de la misma manera que las otras estructuras de rayos X del complejo respiratorio, podemos ver que este electrón se está transportando hacia arriba hacia el espacio intermembrana. Finalmente, este electrón se transfiere a otro hemo, pero este hemo se une en una pequeña proteína móvil, llamada citocromo c. El citocromo c es otro portador de electrones móvil. Lleva el electrón al complejo IV. Volveremos al citocromo c un poco más tarde.
Antes de llegar a eso, ¿a dónde va el otro electrón? Del hemo B L, el segundo electrón se pasa a otro complejo de porfirina de hierro, hemo BH. Eso es hacia la parte inferior de la imagen, así que este electrón en realidad está siendo enviado de vuelta hacia la matriz. Esperando por el hemo B L, sin embargo, hay otra ubiquinona, UQ. Está esperando el electrón. En realidad esperará a dos, luego dejará su muelle y regresará al sitio de unión del ubiquinol (UQH2) para entregar los electrones nuevamente.
¿Por qué molestarse? ¿Por qué no enviar los electrones en su camino correctamente la primera vez? Parece que se están reciclando por alguna razón. Se cree que esta desviación permite bombear protones adicionales a través de la membrana mitocondrial. Cuando la segunda ubiquinona se reduce a ubiquinol a través de la aceptación de dos electrones, también recoge dos protones más de la matriz de abajo. Esos protones eventualmente pasarán hacia el espacio intermembrana cuando el ubiquinol se vuelva a oxidar. Este reciclaje, y exprimir un par de protones adicionales para aumentar el gradiente de protones, se llama bucle Q.
La ventaja del Q-loop se explica en el siguiente diagrama. En el diagrama, las entradas al complejo III se muestran en rojo, las salidas se muestran en azul y los elementos reciclados en verde. Si un ubiquinol simplemente entregara sus electrones y protones y se hiciera, habría dos protones entregados por ubiquinol. Esa es una salida de protones por electrón que fue entrada.
Un segundo ubiquinol haría exactamente lo mismo. Habría cuatro protones de salida para cuatro electrones inicialmente ingresados. Eso sigue siendo una salida de protones por electrón que era entrada.
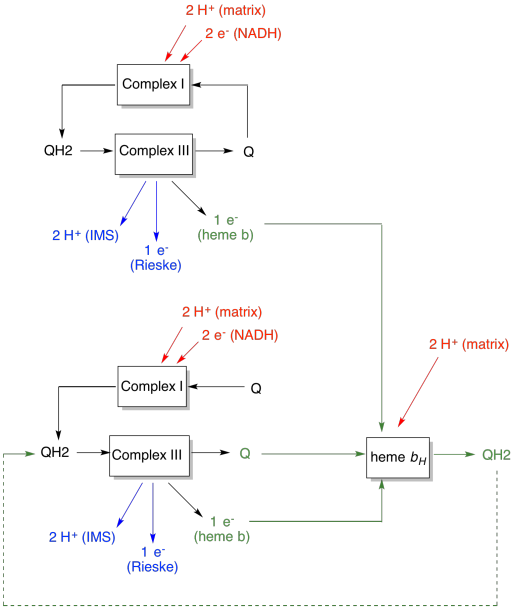
Si, en cambio, se recicla un electrón cada vez, entonces cada segundo ubiquinol conduce a la entrega de un par extra de protones. Eso es porque al recoger los electrones reciclados, una ubiquinona ha tenido que viajar de regreso al lado de la matriz de la membrana y recoger dos protones más. En general, eso significa que se entregan seis protones para la entrada de cuatro electrones, o 1.5 protones de salida por entrada de electrones. Dado que el gradiente de protones es lo que está generando el ATP, entonces al aumentar el número de protones bombeados por electrón que entra, se incrementa la eficiencia.
- El bucle Q aumenta el número de protones bombeados por entrada de electrones al sistema.
Ahora volvamos al portador de electrones saliente, el citocromo c, que transportará los electrones al Complejo IV. En la siguiente imagen, a partir de una estructura cristalina de rayos X diferente, el Complejo se ha encontrado con un acoplamiento de cictocromo c en el sitio de unión. El citocromo c es una proteína globular rosa unida en la parte superior de la imagen, en la mitad izquierda del dímero tal como lo vemos aquí. Está sentado en el borde del espacio intermembrana, que es medio acuoso. A diferencia de la ubiquinona, que se desliza a lo largo de la bicapa lipídica de la membrana, el citocromo c rueda o se desliza o nada sobre la parte superior hasta su destino.
A continuación se muestra la misma vista sin la proteína. La parte inferior de la imagen está bastante abarrotada de lípidos, y hay un azúcar colgando alrededor de la parte superior izquierda, pero justo en la parte superior se puede ver la porfirina de hierro que se encuentra enclavada dentro del citocromo c.
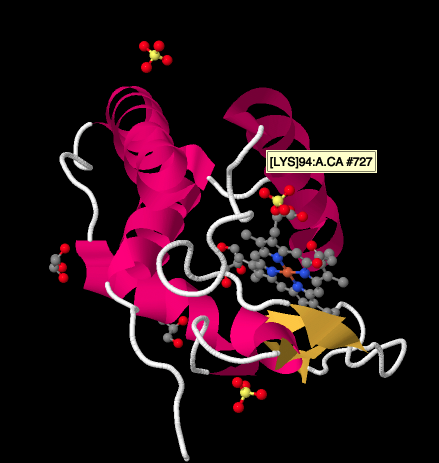
Una mirada más cercana a una estructura de rayos X de un ciclocromo c, a continuación, muestra una imagen relativamente simple en comparación con los complejos que hemos estado viendo. Hay un par de hélices, un par de hojas, algunos bucles, y esa porfirina de hierro o hemo otra vez. Unos pocos iones sulfato están flotando cerca, así como un par de moléculas de glicerol, probablemente introducidas durante la purificación o cristalización de la proteína. Tenga en cuenta que el hemo se encuentra en el borde de la proteína. Esa ubicación puede facilitar la transferencia de electrones desde el Complejo III, o al Complejo IV.
- El citocromo c es el destino final de los electrones que se mueven a través del Complejo III.
- El citocromo c es un portador móvil de un electrón.
- El citocromo c es en realidad una proteína pequeña e hidrófila.
- El citocromo c se encuentra por encima de la membrana mitocondrial, en el borde del espacio intermembranario.
¿Qué impide que el ciclocromo c se desvíe hacia el espacio intermembrana? Si lo hiciera, interrumpiría el flujo eficiente de electrones. Es posible que el citocromo c se mueva de un lado a otro entre el Complejo III y el Complejo IV a través de un mecanismo llamado “dirección”. En la dirección, el portador es guiado a lo largo de una vía a través de cargas complementarias. Por ejemplo, si la membrana está cargada negativamente debido a grupos polares en la superficie de la membrana, entonces las cargas positivas en la superficie del citocromo c pueden restringir su movimiento a la superficie de la membrana. Eso por sí solo reduciría su movilidad de tres dimensiones (en cualquier lugar del espacio intermembrana) a dos dimensiones (en cualquier parte de la superficie de la membrana). Es posible que las interacciones adicionales reduzcan su movilidad aún más a una pista unidimensional entre los dos complejos.
- La dirección, basada en cargas complementarias, evita que el citocromo c se pierda en el espacio intermembrana.
Ejercicio\(\PageIndex{1}\)
Sugerir posibles residuos de aminoácidos en la superficie del citocromo c que pueden ayudar a que permanezca en una superficie de membrana cargada negativamente.
- Responder
-
La arginina y la lisina están cargadas positivamente a pH neutro.
Ejercicio\(\PageIndex{2}\)
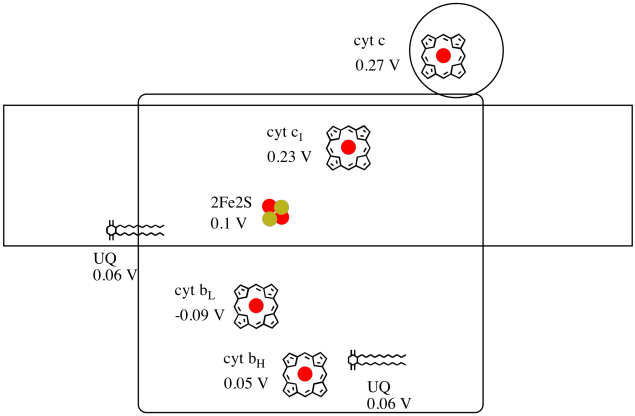
Es difícil medir el potencial de reducción de un sitio individual dentro de una proteína. Sin embargo, los investigadores han podido estimar estos valores midiendo los espectros de EPR bajo diversas condiciones. Suponiendo los potenciales de reducción a continuación, dibuje un diagrama de progreso de reacción para el transporte de un electrón desde el donante inicial de ubiquinona hasta el citocromo c.
- Responder
-
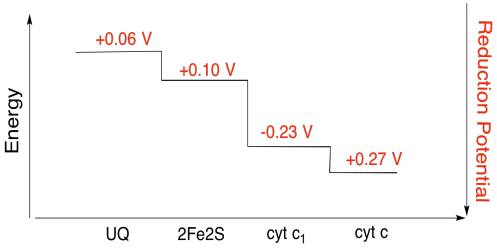
Ejercicio\(\PageIndex{3}\)
Utilizando los valores de la figura anterior, se calcula el cambio de energía cuando se transfiere un electrón del cúmulo de 2Fe2S al citocromo c 1.
- Responder
-
Suponiendo que los potenciales de reducción son:
2Fe2S (buey) + e - → 2Fe2S (rojo) E o rojo = 0.10 V
cyt c 1 (buey) + e - → cyt c 1 (rojo) E o rojo = 0.230 V
Entonces la diferencia de potencial para la reacción, ΔE o = 0.23 - (0.10) V = 0.13 V.
La relación Faraday ΔG = - n F ΔE o da
ΔG = - 1 x 96,485 J V -1 mol -1 x 0.13 V = 12,543 J mol -1 = 12.5 kJ mol -1
Ejercicio\(\PageIndex{4}\)
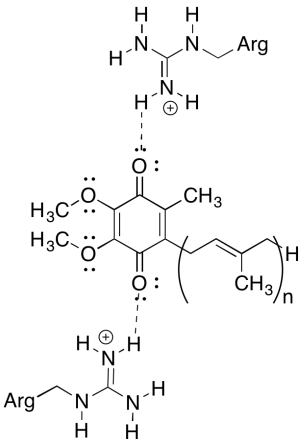
Una de las cosas desplejas de este complejo es que, en el bucle de reciclaje de electrones, la transferencia de electrones parece pasar de una ubiquinona inicial a una ubiquinona final, sin fuerza motriz general. Sin embargo, los potenciales de reducción son muy sensibles al medio ambiente. Mostrar cómo los residuos de arginina cercanos podrían hacer que el potencial de reducción de una ubiquinona sea más positivo.
- Responder
-
Los residuos de arginina positivos conferirían carga positiva parcial a la ubiquinona a través de enlaces de hidrógeno; la ubiquinona tendría un potencial de reducción más positivo como resultado.
Consulte la página de enfoque sobre la estructura del Complejo III en Biochemistry Online.
Estructura cristalina de rayos X del Complejo III de: Gao, X., Wen, X., Esser, L., Quinn, B., Yu, L., Yu, C.-A., Xia, D. Bases estructurales para la reducción de quinona en el complejo bc1: un análisis comparativo de las estructuras cristalinas del citocromo mitocondrial bc1 con sustrato unido e inhibidores en el sitio Qi. Bioquímica 2003, 4, 9067-9080. (1NTZ)
Estructura cristalina de rayos X del Complejo III con citocromo unido c de: Solmaz, S.R., Hunte, C. Estructura del complejo III con citocromo unido c en estado reducido y definición de una interfaz central mínima para transferencia electrónica. J. Biol. Chem. 2008, 283, 17542-17549. (3CXH)
Estructura cristalina de rayos X del citocromo c de: Enguita, F.J., Pohl, E., Turner, D.L., Santos, H., Carrondo, M.A. Evidencia estructural de una vía de transferencia de protones acoplada a la reducción del heno del citocromo c” de Methylophilus methylotrophus. J. Biol. Inorg. Chem. 2006 11, 189. (1OAE)