
21.4: La Respuesta Inmune Adaptativa - Linfocitos B y Anticuerpos
- Page ID
- 123216

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Explicar cómo maduran las células B y cómo se desarrolla la tolerancia de células B
- Discutir cómo se activan las células B y se diferencian en células plasmáticas
- Describir la estructura de las clases de anticuerpos y sus funciones
Los anticuerpos fueron el primer componente de la respuesta inmune adaptativa que caracterizaron los científicos que trabajan en el sistema inmune. Ya se sabía que los individuos que sobrevivieron a una infección bacteriana eran inmunes a la reinfección con el mismo patógeno. Los primeros microbiólogos tomaron suero de un paciente inmune y lo mezclaron con un cultivo fresco del mismo tipo de bacteria, luego observaron la bacteria bajo un microscopio. Las bacterias se agruparon en un proceso llamado aglutinación. Cuando se utilizó una especie bacteriana diferente, la aglutinación no ocurrió. Así, había algo en el suero de los individuos inmunes que se podían unir específicamente a las bacterias y aglutinar.
Los científicos ahora saben que la causa de la aglutinación es una molécula de anticuerpo, también llamada inmunoglobulina. ¿Qué es un anticuerpo? Una proteína de anticuerpo es esencialmente una forma secretada de un receptor de células B. (De hecho, la inmunoglobulina de superficie es otro nombre para el receptor de células B). No es sorprendente que los mismos genes codifican tanto los anticuerpos secretados como las inmunoglobulinas de superficie. Una diferencia menor en la forma en que se sintetizan estas proteínas distingue una célula B virgen con anticuerpo en su superficie de una célula plasmática secretora de anticuerpos sin anticuerpos en su superficie. Los anticuerpos de la célula plasmática tienen exactamente el mismo sitio de unión a antígeno y especificidad que sus precursores de células B.
Hay cinco clases diferentes de anticuerpos encontrados en humanos: IgM, IgD, IgG, IgA e IgE. Cada uno de estos tiene funciones específicas en la respuesta inmune, por lo que al aprender sobre ellas, los investigadores pueden conocer la gran variedad de funciones de anticuerpos críticas para muchas respuestas inmunes adaptativas.
Las células B no reconocen el antígeno en la forma compleja de las células T. Las células B pueden reconocer antígeno nativo sin procesar y no requieren la participación de moléculas MHC y células presentadoras de antígeno.
Diferenciación y Activación de Células B
Las células B se diferencian en la médula ósea. Durante el proceso de maduración, se generan hasta 100 billones de clones diferentes de células B, lo que es similar a la diversidad de receptores de antígeno que se observan en las células T.
La diferenciación de células B y el desarrollo de tolerancia no se entienden tan bien como en las células T. La tolerancia central es la destrucción o inactivación de células B que reconocen autoantígenos en la médula ósea, y su papel es crítico y bien establecido. En el proceso de deleción clonal, las células B inmaduras que se unen fuertemente a autoantígenos expresados en los tejidos son señaladas para suicidarse por apoptosis, eliminándolas de la población. En el proceso de anergia clonal, sin embargo, las células B expuestas a antígeno soluble en la médula ósea no se eliminan físicamente, sino que se vuelven incapaces de funcionar.
Otro mecanismo llamado tolerancia periférica es un resultado directo de la tolerancia de células T. En tolerancia periférica, las células B maduras funcionales abandonan la médula ósea pero aún no se han expuesto al autoantígeno. La mayoría de los antígenos proteicos requieren señales de las células T colaboradoras (Th2) para proceder a producir anticuerpos. Cuando una célula B se une a un autoantígeno pero no recibe señales de una célula Th2 cercana para producir anticuerpos, se señala que la célula sufre apoptosis y se destruye. Este es otro ejemplo más del control que tienen las células T sobre la respuesta inmune adaptativa.
Después de que las células B se activan por su unión al antígeno, se diferencian en células plasmáticas. Las células plasmáticas suelen abandonar los órganos linfoides secundarios, donde se genera la respuesta, y vuelven a migrar a la médula ósea, donde comenzó todo el proceso de diferenciación. Después de secretar anticuerpos por un periodo específico, mueren, ya que la mayor parte de su energía se dedica a fabricar anticuerpos y no a mantenerse. Así, se dice que las células plasmáticas están diferenciadas terminalmente.
La célula B final de interés es la célula B de memoria, que resulta de la expansión clonal de una célula B activada. Las células B de memoria funcionan de manera similar a las células T de memoria. Conllevan a una respuesta secundaria más fuerte y rápida en comparación con la respuesta primaria, como se ilustra a continuación.
Estructura de anticuerpos
Los anticuerpos son glicoproteínas que consisten en dos tipos de cadenas polipeptídicas con carbohidratos unidos. La cadena pesada y la cadena ligera son los dos polipéptidos que forman el anticuerpo. Las principales diferencias entre las clases de anticuerpos están en las diferencias entre sus cadenas pesadas, pero como verá, las cadenas ligeras tienen un papel importante, formando parte del sitio de unión al antígeno en las moléculas del anticuerpo.
Modelos de cuatro cadenas de estructuras de anticuerpos
Todas las moléculas de anticuerpo tienen dos cadenas pesadas idénticas y dos cadenas ligeras idénticas. (Algunos anticuerpos contienen múltiples unidades de esta estructura de cuatro cadenas). La región Fc del anticuerpo está formada por las dos cadenas pesadas que se unen, generalmente unidas por enlaces disulfuro (Figura\(\PageIndex{1}\)). La porción Fc del anticuerpo es importante ya que muchas células efectoras del sistema inmune tienen receptores Fc. Las células que tienen estos receptores pueden unirse entonces a patógenos recubiertos con anticuerpos, aumentando enormemente la especificidad de las células efectoras. En el otro extremo de la molécula hay dos sitios de unión a antígeno idénticos.

Cinco clases de anticuerpos y sus funciones
En general, los anticuerpos tienen dos funciones básicas. Pueden actuar como receptores de antígeno de células B o pueden ser secretados, circulados y unirse a un patógeno, a menudo marcándolo para su identificación por otras formas de respuesta inmune. De las cinco clases de anticuerpos, observe que solo dos pueden funcionar como receptor de antígeno para las células B vínicas: IgM e IgD (Figura\(\PageIndex{2}\)). Las células B maduras que salen de la médula ósea expresan tanto IgM como IgD, pero ambos anticuerpos tienen la misma especificidad antigénica. Sin embargo, solo se secreta IgM y no se ha descubierto ninguna otra función no receptora para IgD.

IgM consiste en cinco estructuras de cuatro cadenas (20 cadenas totales con 10 sitios de unión a antígeno idénticos) y, por lo tanto, es la más grande de las moléculas de anticuerpo. IgM suele ser el primer anticuerpo producido durante una respuesta primaria. Sus 10 sitios de unión al antígeno y su gran forma le permiten unirse bien a muchas superficies bacterianas. Es excelente para unir proteínas del complemento y activar la cascada del complemento, consistente con su papel en la promoción de la quimiotaxis, opsonización y lisis celular. Por lo tanto, es un anticuerpo muy efectivo contra bacterias en etapas tempranas de una respuesta de anticuerpos primarios. A medida que avanza la respuesta primaria, el anticuerpo producido en una célula B puede cambiar a IgG, IgA o IgE mediante el proceso conocido como cambio de clase. El cambio de clase es el cambio de una clase de anticuerpo a otra. Si bien la clase de anticuerpos cambia, la especificidad y los sitios de unión al antígeno no lo hacen. Así, los anticuerpos elaborados siguen siendo específicos del patógeno que estimuló la respuesta inicial de IgM.
La IgG es un anticuerpo principal de respuestas primarias tardías y el principal anticuerpo de respuestas secundarias en la sangre. Esto se debe a que el cambio de clase ocurre durante las respuestas primarias. La IgG es un anticuerpo monomérico que elimina patógenos de la sangre y puede activar proteínas del complemento (aunque no tan bien como IgM), aprovechando sus actividades antibacterianas. Además, esta clase de anticuerpos es el que cruza la placenta para proteger al feto en desarrollo de la enfermedad que sale de la sangre al líquido intersticial para combatir los patógenos extracelulares.
La IgA existe en dos formas, un monómero de cuatro cadenas en la sangre y una estructura de ocho cadenas, o dímero, en las secreciones de glándulas exocrinas de las membranas mucosas, incluyendo moco, saliva y lágrimas. Por lo tanto, la IgA dimérica es el único anticuerpo que abandona el interior del cuerpo para proteger las superficies corporales. La IgA también es de importancia para los recién nacidos, ya que este anticuerpo está presente en la leche materna (calostro), que sirve para proteger al lactante de enfermedades.
La IgE suele asociarse con alergias y anafilaxia. Está presente en la concentración más baja en la sangre, debido a que su región Fc se une fuertemente a un receptor Fc específico de IgE en las superficies de los mastocitos. La IgE hace que la desgranulación de mastocitos sea muy específica, de tal manera que si una persona es alérgica al maní, habrá IgE específica de cacahuete unida a sus mastocitos. En esta persona, comer cacahuetes provocará que los mastocitos se desgranulen, ocasionando en ocasiones reacciones alérgicas graves, incluyendo anafilaxia, una respuesta alérgica severa y sistémica que puede causar la muerte.
Selección Clonal de Células B
La selección y expansión clonal funcionan de la misma manera en las células B que en las células T. Solo se seleccionan y expanden las células B con especificidad antigénica apropiada (Figura\(\PageIndex{3}\)). Finalmente, las células plasmáticas secretan anticuerpos con especificidad antigénica idéntica a los que estaban en las superficies de las células B seleccionadas. Observe en la figura que tanto las células plasmáticas como las células B de memoria se generan simultáneamente.
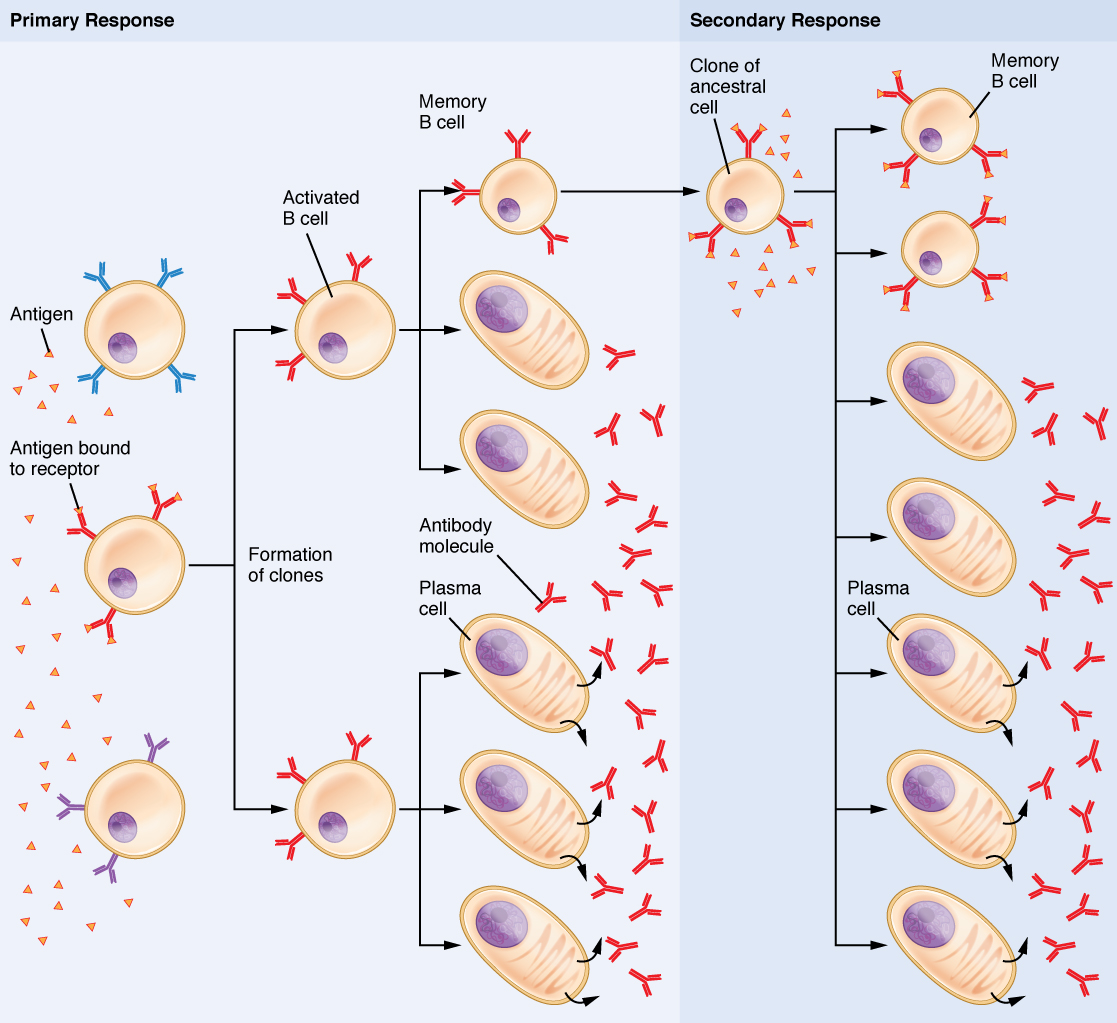
Respuestas de células B primarias versus secundarias
Las respuestas primarias y secundarias en relación con las células T se discutieron anteriormente. En esta sección se analizarán estas respuestas con células B y producción de anticuerpos. Debido a que los anticuerpos se obtienen fácilmente de muestras de sangre, son fáciles de seguir y graficar (Figura\(\PageIndex{4}\)). Como verá en la figura, la respuesta primaria a un antígeno (que representa un patógeno) se retrasa varios días. Este es el tiempo que tardan los clones de células B en expandirse y diferenciarse en células plasmáticas. El nivel de anticuerpos producidos es bajo, pero es suficiente para la protección inmune. La segunda vez que una persona se encuentra con el mismo antígeno, no hay retraso de tiempo, y la cantidad de anticuerpo que se produce es mucho mayor. Así, la respuesta secundaria de anticuerpos abruma rápidamente a los patógenos y, en la mayoría de las situaciones, no se sienten síntomas. Cuando se usa un antígeno diferente, se realiza otra respuesta primaria con sus bajos niveles de anticuerpos y retraso de tiempo.

Inmunidad activa versus pasiva
La inmunidad a los patógenos, y la capacidad de controlar el crecimiento de patógenos para que el daño a los tejidos del cuerpo sea limitado, puede ser adquirida por (1) el desarrollo activo de una respuesta inmune en el individuo infectado o (2) la transferencia pasiva de componentes inmunes de un individuo inmune a uno no inmune. Tanto la inmunidad activa como la pasiva tienen ejemplos en el mundo natural y como parte de la medicina.
La inmunidad activa es la resistencia a patógenos adquiridos durante una respuesta inmune adaptativa dentro de un individuo (Tabla). La inmunidad activa adquirida naturalmente, la respuesta a un patógeno, es el foco de este capítulo. La inmunidad activa adquirida artificialmente implica el uso de vacunas. Una vacuna es un patógeno muerto o debilitado o sus componentes que, cuando se administra a un individuo sano, conduce al desarrollo de la memoria inmunológica (una respuesta inmune primaria debilitada) sin causar mucho en el camino de los síntomas. Así, con el uso de vacunas, se puede evitar el daño por enfermedad que resulta de la primera exposición al patógeno, sin embargo, cosechar los beneficios de la protección contra la memoria inmunológica. El advenimiento de las vacunas fue uno de los mayores avances médicos del siglo XX y condujo a la erradicación de la viruela y al control de muchas enfermedades infecciosas, entre ellas la poliomielitis, el sarampión y la tos ferina.
| Inmunidad activa versus pasiva | ||
|---|---|---|
| Natural | Artificial | |
| Activo | Respuesta inmune adaptativa | Respuesta vacunal |
| Pasivo | Anticuerpos transplacentarios/lactancia | Inyecciones de inmunoglobulina |
La inmunidad pasiva surge de la transferencia de anticuerpos a un individuo sin requerir que monten su propia respuesta inmune activa. La inmunidad pasiva adquirida de forma natural se observa durante el desarrollo fetal. La IgG se transfiere de la circulación materna al feto a través de la placenta, protegiendo al feto de la infección y protegiendo al recién nacido durante los primeros meses de su vida. Como ya se dijo, un recién nacido se beneficia de los anticuerpos IgA que obtiene de la leche durante la lactancia. El feto y el recién nacido se benefician así de la memoria inmunológica de la madre a los patógenos a los que ha estado expuesta. En medicina, la inmunidad pasiva adquirida artificialmente generalmente implica inyecciones de inmunoglobulinas, tomadas de animales previamente expuestos a un patógeno específico. Este tratamiento es un método de acción rápida para proteger temporalmente a un individuo que posiblemente estuvo expuesto a un patógeno. La desventaja de ambos tipos de inmunidad pasiva es la falta del desarrollo de la memoria inmunológica. Una vez que se transfieren los anticuerpos, son efectivos solo por un tiempo limitado antes de que se degraden.

La inmunidad se puede adquirir de forma activa o pasiva, y puede ser natural o artificial. Mire este video para ver una discusión animada sobre la inmunidad pasiva y activa. ¿Cuál es un ejemplo de inmunidad natural adquirida pasivamente?
Antígenos dependientes de células T versus independientes de células T
Como se discutió anteriormente, las células Th2 secretan citocinas que impulsan la producción de anticuerpos en una célula B, respondiendo a antígenos complejos como los producidos por las proteínas. Por otro lado, algunos antígenos son independientes de las células T. Un antígeno independiente de células T generalmente está en forma de restos de carbohidratos repetidos que se encuentran en las paredes celulares de las bacterias. Cada anticuerpo en la superficie de las células B tiene dos sitios de unión, y la naturaleza repetida del antígeno independiente de las células T conduce a la reticulación de los anticuerpos de superficie en la célula B. El entrecruzamiento es suficiente para activarlo en ausencia de citocinas de células T.
Un antígeno dependiente de células T, por otro lado, generalmente no se repite en el mismo grado sobre el patógeno y por lo tanto no reticula el anticuerpo de superficie con la misma eficiencia. Para provocar una respuesta a dichos antígenos, las células B y T deben acercarse entre sí (Figura\(\PageIndex{5}\)). La célula B debe recibir dos señales para activarse. Su inmunoglobulina de superficie debe reconocer antígeno nativo. Parte de este antígeno se internaliza, procesa y presenta a las células Th2 en una molécula MHC clase II. La célula T luego se une usando su receptor de antígeno y se activa para secretar citocinas que se difunden a la célula B, activándola finalmente por completo. Así, la célula B recibe señales tanto de su anticuerpo de superficie como de la célula T a través de sus citocinas, y actúa como una célula presentadora de antígeno profesional en el proceso.

Revisión del Capítulo
Las células B, que se desarrollan dentro de la médula ósea, son las encargadas de elaborar cinco clases diferentes de anticuerpos, cada uno con sus propias funciones. Las células B tienen sus propios mecanismos de tolerancia, pero en la tolerancia periférica, las células B que salen de la médula ósea permanecen inactivas debido a la tolerancia de las células T. Algunas células B no necesitan citocinas de células T para producir anticuerpos, y evitan esta necesidad por la reticulación de su inmunoglobulina de superficie por residuos repetidos de carbohidratos que se encuentran en las paredes celulares de muchas especies bacterianas. Otros requieren que las células T se activen.
Preguntas de Enlace Interactivo
La inmunidad se puede adquirir de forma activa o pasiva, y puede ser natural o artificial. Mire este video para ver una discusión animada sobre la inmunidad pasiva y activa. ¿Cuál es un ejemplo de inmunidad natural adquirida pasivamente?
Respuesta: La lactancia materna es un ejemplo de inmunidad natural adquirida pasivamente.
Capítulo de revisión
P. Eliminar la funcionalidad de una célula B sin matarla se llama ________.
A. selección clonal
B. expansión clonal
C. deleción clonal
D. anería clonal
Respuesta: D
P. ¿Qué clase de anticuerpos cruza la placenta en mujeres embarazadas?
A. IgM
B. IgA
C. IgE
D. IgG
Respuesta: D
P. ¿Qué clase de anticuerpo no tiene otra función conocida que no sea como receptor de antígeno?
A. IgM
B. IgA
C. IgE
D. IgD
Respuesta: D
P: ¿Cuándo ocurre el cambio de clase?
A. respuesta primaria
B. respuesta secundaria
C. tolerancia
D. respuesta de memoria
Respuesta: A
P. ¿Qué clase de anticuerpo se encuentra en la mucosidad?
A. IgM
B. IgA
C. IgE
D. IgD
Respuesta: B
Preguntas de Pensamiento Crítico
P. Describir cómo se desarrollan las respuestas secundarias de células B.
A. Las células B activadas durante una respuesta primaria se diferencian en células plasmáticas diferenciadas terminalmente o en células B de memoria. Estas células B de memoria son las que responden durante una respuesta secundaria o de anticuerpos de memoria.
P. Describir el papel de la IgM en la inmunidad.
A. IgM es un receptor de antígeno en células B sin tratamiento previo. Tras la activación, las células B vínicas producen IgM primero. IgM es bueno para unirse al complemento y, por lo tanto, tiene buenos efectos antibacterianos. IgM se reemplaza con otras clases de anticuerpos más adelante en la respuesta primaria debido al cambio de clase.
Glosario
- inmunidad activa
- inmunidad desarrollada a partir del propio sistema inmunológico de un individuo
- tolerancia central
- Tolerancia de células B inducida en células B inmaduras de la médula ósea
- conmutación de clase
- capacidad de las células B para cambiar la clase de anticuerpo que producen sin alterar la especificidad para el antígeno
- anería clonal
- proceso mediante el cual las células B que reaccionan a antígenos solubles en la médula ósea se vuelven no funcionales
- deleción clonal
- eliminación de células B autorreactivas mediante la inducción de apoptosis
- Región Fc
- en una molécula de anticuerpo, el sitio donde se unen los dos términos de las cadenas pesadas; muchas células tienen receptores para esta porción del anticuerpo, agregando funcionalidad a estas moléculas
- cadena pesada
- cadena proteica más grande de un anticuerpo
- IgA
- anticuerpo cuyo dímero es secretado por las glándulas exocrinas, es especialmente efectivo contra patógenos digestivos y respiratorios, y puede transmitir inmunidad a un lactante a través de la lactancia
- IgD
- clase de anticuerpo cuya única función conocida es como receptor en células B sin tratamiento previo; importante en la activación de células B
- IgE
- anticuerpo que se une a mastocitos y causa desgranulación específica de antígeno durante una respuesta alérgica
- IgG
- principal anticuerpo sanguíneo de respuestas primarias tardías y secundarias tempranas; pasó de la madre al feto vía placenta
- IgM
- anticuerpo cuyo monómero es un receptor superficial de células B vívidas; el pentámero es el primer anticuerpo hecho plasma sanguíneo durante las respuestas primarias
- inmunoglobulina
- anticuerpo proteico; ocurre como una de las cinco clases principales
- cadena ligera
- cadena proteica pequeña de un anticuerpo
- inmunidad pasiva
- transferencia de inmunidad a un patógeno a un individuo que carece de inmunidad a este patógeno generalmente mediante la inyección de anticuerpos
- tolerancia periférica
- células B maduras hechas tolerantes por falta de ayuda de células T
- Antígeno dependiente de células T
- antígeno que se une a las células B, que requiere señales de las células T para producir anticuerpos
- Antígeno independiente de células T
- se une a las células B, que no requieren señales de las células T para producir anticuerpos