
24.2: Metabolismo de los carbohidratos
- Page ID
- 122918

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Explicar los procesos de glucólisis
- Describir la ruta de una molécula de piruvato a través del ciclo de Krebs
- Explicar el transporte de electrones a través de la cadena de transporte de electrones
- Describir el proceso de producción de ATP mediante fosforilación oxidativa
- Resumir el proceso de gluconeogénesis
Los carbohidratos son moléculas orgánicas compuestas por átomos de carbono, hidrógeno y oxígeno. La familia de carbohidratos incluye azúcares tanto simples como complejos. La glucosa y la fructosa son ejemplos de azúcares simples, y el almidón, el glucógeno y la celulosa son ejemplos de azúcares complejos. Los azúcares complejos también se llaman polisacáridos y están hechos de múltiples moléculas de monosacáridos. Los polisacáridos sirven como almacenamiento de energía (por ejemplo, almidón y glucógeno) y como componentes estructurales (por ejemplo, quitina en insectos y celulosa en plantas).
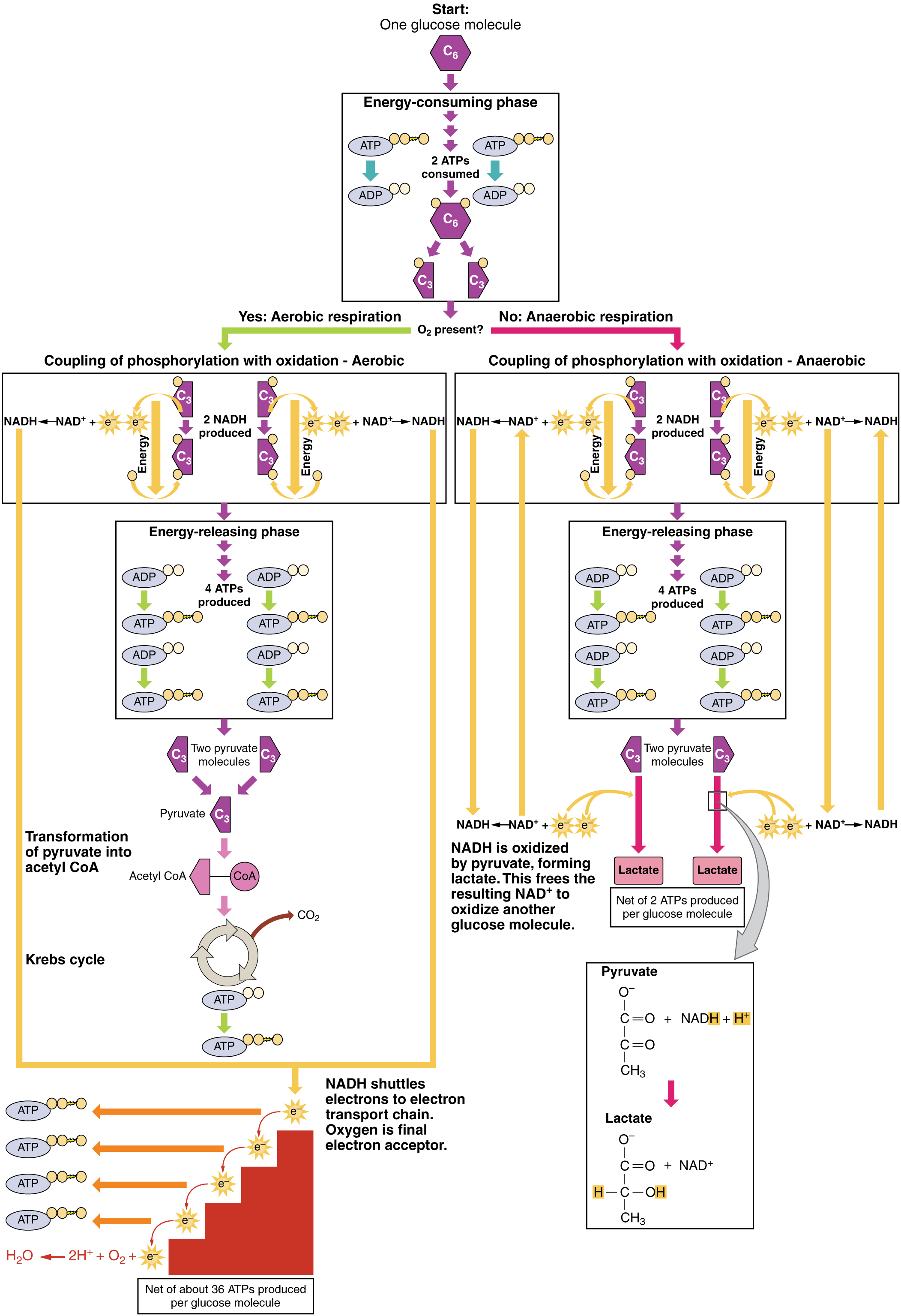
Durante la digestión, los carbohidratos se descomponen en azúcares simples y solubles que pueden transportarse a través de la pared intestinal hacia el sistema circulatorio para ser transportados por todo el cuerpo. La digestión de carbohidratos comienza en la boca con la acción de la amilasa salival sobre los almidones y termina con monosacáridos que se absorben a través del epitelio del intestino delgado. Una vez que los monosacáridos absorbidos son transportados a los tejidos, comienza el proceso de respiración celular (Figura\(\PageIndex{1}\)). Esta sección se centrará primero en la glucólisis, un proceso donde se oxida la glucosa monosacárido, liberando la energía almacenada en sus enlaces para producir ATP.

Glicólisis
La glucosa es la fuente de energía más fácilmente disponible en el cuerpo. Después de que los procesos digestivos descomponen los polisacáridos en monosacáridos, incluida la glucosa, los monosacáridos se transportan a través de la pared del intestino delgado y al sistema circulatorio, que los transporta al hígado. En el hígado, los hepatocitos pasan la glucosa a través del sistema circulatorio o almacenan el exceso de glucosa como glucógeno. Las células del cuerpo absorben la glucosa circulante en respuesta a la insulina y, a través de una serie de reacciones llamadas glucólisis, transfieren parte de la energía en la glucosa a ADP para formar ATP (Figura\(\PageIndex{2}\)). El último paso en la glucólisis produce el producto piruvato.
La glucólisis comienza con la fosforilación de glucosa por hexoquinasa para formar glucosa-6-fosfato. En este paso se utiliza un ATP, que es el donante del grupo fosfato. Bajo la acción de la fosfofructoquinasa, la glucosa-6-fosfato se convierte en fructosa-6-fosfato. En este punto, un segundo ATP dona su grupo fosfato, formando fructosa-1,6-bisfosfato. Este azúcar de seis carbonos se divide para formar dos moléculas fosforiladas de tres carbonos, gliceraldehído-3-fosfato y dihidroxiacetona fosfato, las cuales se convierten ambas en gliceraldehído-3-fosfato. El gliceraldehído-3-fosfato se fosforila adicionalmente con grupos donados por dihidrógeno fosfato presentes en la célula para formar la molécula de tres carbonos 1,3-bisfosfosglicerato. La energía de esta reacción proviene de la oxidación de (eliminación de electrones de) gliceraldehído-3-fosfato. En una serie de reacciones que conducen al piruvato, los dos grupos fosfato se transfieren luego a dos ADP para formar dos ATP. Así, la glucólisis utiliza dos ATPs pero genera cuatro ATPs, produciendo una ganancia neta de dos ATPs y dos moléculas de piruvato. En presencia de oxígeno, el piruvato continúa hasta el ciclo de Krebs (también llamado ciclo del ácido cítrico o ciclo del ácido tricarboxílico (TCA), donde se extrae energía adicional y se transmite.


La glucólisis se puede dividir en dos fases: consumo de energía (también llamado cebado químico) y rendimiento energético. La primera fase es la fase de consumo de energía, por lo que requiere de dos moléculas de ATP para iniciar la reacción por cada molécula de glucosa. Sin embargo, el final de la reacción produce cuatro ATP, lo que resulta en una ganancia neta de dos moléculas de energía ATP.
La glucólisis se puede expresar como la siguiente ecuación:
Glucosa + 2ATP + 2NAD + + 4ADP + 2P i → 2 Piruvato + 4ATP + 2NADH + 2H +
Esta ecuación establece que la glucosa, en combinación con ATP (la fuente de energía), NAD + (una coenzima que sirve como aceptor de electrones) y fosfato inorgánico, se descompone en dos moléculas de piruvato, generando cuatro moléculas de ATP, para un rendimiento neto de dos ATP y dos coenzimas NADH que contienen energía. El NADH que se produce en este proceso se utilizará posteriormente para producir ATP en las mitocondrias. Es importante destacar que al final de este proceso, una molécula de glucosa genera dos moléculas de piruvato, dos moléculas de ATP de alta energía y dos moléculas de NADH portadoras de electrones.
Las siguientes discusiones sobre la glucólisis incluyen las enzimas responsables de las reacciones. Cuando la glucosa ingresa a una célula, la enzima hexoquinasa (o glucoquinasa, en el hígado) agrega rápidamente un fosfato para convertirlo en glucosa-6-fosfato. Una quinasa es un tipo de enzima que agrega una molécula de fosfato a un sustrato (en este caso, glucosa, pero también puede ser cierto para otras moléculas). Esta etapa de conversión requiere un ATP y esencialmente atrapa la glucosa en la célula, evitando que vuelva a pasar a través de la membrana plasmática, permitiendo así que proceda la glucólisis. También funciona para mantener un gradiente de concentración con niveles de glucosa más altos en la sangre que en los tejidos. Al establecer este gradiente de concentración, la glucosa en la sangre podrá fluir de un área de alta concentración (la sangre) a una zona de baja concentración (los tejidos) para ser utilizada o almacenada. La hexoquinasa se encuentra en casi todos los tejidos del cuerpo. La glucoquinasa, por otro lado, se expresa en tejidos que son activos cuando los niveles de glucosa en sangre son altos, como el hígado. La hexoquinasa tiene una mayor afinidad por la glucosa que la glucoquinasa y por lo tanto es capaz de convertir la glucosa a un ritmo más rápido que la glucoquinasa. Esto es importante cuando los niveles de glucosa son muy bajos en el organismo, ya que permite que la glucosa viaje preferentemente a aquellos tejidos que más la requieren.
En el siguiente paso de la primera fase de la glucólisis, la enzima glucosa-6-fosfato isomerasa convierte la glucosa-6-fosfato en fructosa-6-fosfato. Al igual que la glucosa, la fructosa es también un azúcar que contiene seis carbonos. La enzima fosfofructoquinasa-1 luego agrega un fosfato más para convertir la fructosa-6-fosfato en fructosa-1-6-bisfosfato, otro azúcar de seis carbonos, usando otra molécula de ATP. La aldolasa luego descompone este fructosa-1-6-bisfosfato en dos moléculas de tres carbonos, gliceraldehído-3-fosfato y dihidroxiacetona fosfato. La enzima triosafosfato isomerasa convierte el fosfato de dihidroxiacetona en una segunda molécula de gliceraldehído-3-fosfato. Por lo tanto, al final de esta fase de cebado químico o de consumo de energía, una molécula de glucosa se descompone en dos moléculas de gliceraldehído-3-fosfato.
La segunda fase de la glucólisis, la fase de producción de energía, crea la energía que es el producto de la glucólisis. La gliceraldehído-3-fosfato deshidrogenasa convierte cada gliceraldehído-3-fosfato de tres carbonos producido durante la fase de consumo de energía en 1,3-bisfosfosglicerato. Esta reacción libera un electrón que luego es captado por NAD + para crear una molécula de NADH. El NADH es una molécula de alta energía, como el ATP, pero a diferencia del ATP, no es utilizada como moneda de energía por la célula. Debido a que hay dos moléculas de gliceraldehído-3-fosfato, durante esta etapa se sintetizan dos moléculas de NADH. Cada 1,3-bisfosfosglicerato se desfosforila posteriormente (es decir, se elimina un fosfato) por fosfoglicerato quinasa en 3-fosfoglicerato. Cada fosfato liberado en esta reacción puede convertir una molécula de ADP en una molécula de ATP de alta energía, lo que resulta en una ganancia de dos moléculas de ATP.
La enzima fosfoglicerato mutasa luego convierte las moléculas de 3-fosfoglicerato en 2-fosfoglicerato. La enzima enolasa actúa entonces sobre las moléculas de 2-fosfoglicerato para convertirlas en moléculas de fosfoenolpiruvato. El último paso de la glucólisis implica la desfosforilación de las dos moléculas de fosfoenolpiruvato por la piruvato quinasa para crear dos moléculas de piruvato y dos moléculas de ATP.
En resumen, una molécula de glucosa se descompone en dos moléculas de piruvato y crea dos moléculas netas de ATP y dos moléculas de NADH por glucólisis. Por lo tanto, la glucólisis genera energía para la célula y crea moléculas de piruvato que pueden procesarse más a través del ciclo aeróbico de Krebs (también llamado ciclo del ácido cítrico o ciclo del ácido tricarboxílico); convertirse en ácido láctico o alcohol (en levadura) por fermentación; o usarse posteriormente para la síntesis de glucosa a través de la gluconeogénesis.
Respiración anaeróbica
Cuando el oxígeno es limitado o ausente, el piruvato entra en una vía anaeróbica. En estas reacciones, el piruvato se puede convertir en ácido láctico. Además de generar un ATP adicional, esta vía sirve para mantener baja la concentración de piruvato para que la glucólisis continúe, y oxida el NADH en el NAD + que necesita la glucólisis. En esta reacción, el ácido láctico reemplaza al oxígeno como aceptor final de electrones. La respiración anaeróbica ocurre en la mayoría de las células del cuerpo cuando el oxígeno es limitado o las mitocondrias están ausentes o no son funcionales. Por ejemplo, debido a que los eritrocitos (glóbulos rojos) carecen de mitocondrias, deben producir su ATP a partir de la respiración anaeróbica. Esta es una vía efectiva de producción de ATP por cortos periodos de tiempo, que van desde segundos hasta unos minutos. El ácido láctico producido se difunde al plasma y se transporta al hígado, donde se vuelve a convertir en piruvato o glucosa a través del ciclo de Cori. De igual manera, cuando una persona hace ejercicio, los músculos usan ATP más rápido de lo que se le puede entregar oxígeno. Dependen de la glucólisis y la producción de ácido láctico para una rápida producción de ATP.
Respiración aeróbica
En presencia de oxígeno, el piruvato puede ingresar al ciclo de Krebs donde se extrae energía adicional a medida que los electrones se transfieren del piruvato a los receptores NAD +, GDP y FAD, siendo el dióxido de carbono un “producto de desecho” (Figura\(\PageIndex{3}\)). El NADH y el FADH 2 pasan electrones a la cadena de transporte de electrones, que utiliza la energía transferida para producir ATP. Como paso terminal en la cadena de transporte de electrones, el oxígeno es el aceptor de electrones terminal y crea agua dentro de las mitocondrias.
Ciclo de Krebs/Ciclo de ácido cítrico/Ciclo de ácido tricarboxílico
Las moléculas de piruvato generadas durante la glucólisis son transportadas a través de la membrana mitocondrial hacia la matriz mitocondrial interna, donde son metabolizadas por enzimas en una vía llamada ciclo de Krebs (Figura\(\PageIndex{4}\)). El ciclo de Krebs también se llama comúnmente el ciclo del ácido cítrico o el ciclo del ácido tricarboxílico (TCA). Durante el ciclo de Krebs, se crean moléculas de alta energía, incluyendo ATP, NADH y FADH 2. NADH y FADH 2 luego pasan electrones a través de la cadena de transporte de electrones en las mitocondrias para generar más moléculas de ATP.


La molécula de piruvato de tres carbonos generada durante la glucólisis se mueve del citoplasma a la matriz mitocondrial, donde es convertida por la enzima piruvato deshidrogenasa en una molécula de acetil coenzima A (acetil CoA) de dos carbonos. Esta reacción es una reacción de descarboxilación oxidativa. Convierte el piruvato de tres carbonos en una molécula de acetil CoA de dos carbonos, liberando dióxido de carbono y transfiriendo dos electrones que se combinan con NAD + para formar NADH. El acetil CoA entra en el ciclo de Krebs combinándolo con una molécula de cuatro carbonos, el oxaloacetato, para formar la molécula citrato de seis carbonos, o ácido cítrico, al mismo tiempo que libera la molécula de coenzima A.
La molécula de citrato de seis carbonos se convierte sistemáticamente en una molécula de cinco carbonos y luego en una molécula de cuatro carbonos, terminando con oxaloacetato, el inicio del ciclo. En el camino, cada molécula de citrato producirá un ATP, un FADH 2 y tres NADH. El FADH 2 y el NADH ingresarán al sistema de fosforilación oxidativa ubicado en la membrana mitocondrial interna. Además, el ciclo de Krebs suministra los materiales de partida para procesar y descomponer proteínas y grasas.
Para iniciar el ciclo de Krebs, la citrato sintasa combina acetil CoA y oxaloacetato para formar una molécula de citrato de seis carbonos; posteriormente se libera CoA y puede combinarse con otra molécula de piruvato para comenzar el ciclo nuevamente. La enzima aconitasa convierte el citrato en isocitrato. En dos etapas sucesivas de descarboxilación oxidativa, se producen dos moléculas de CO 2 y dos moléculas de NADH cuando la isocitrato deshidrogenasa convierte el isocitrato en el α-cetoglutarato de cinco carbonos, que luego es catalizado y convertido en el succinilo CoA de cuatro carbonos por α-cetoglutarato deshidrogenasa. La enzima succinilCoA deshidrogenasa luego convierte succinyl CoA en succinato y forma la molécula de alta energía GTP, que transfiere su energía a ADP para producir ATP. La succinato deshidrogenasa luego convierte el succinato en fumarato, formando una molécula de FADH 2. La fumarasa luego convierte el fumarato en malato, que la malato deshidrogenasa luego convierte nuevamente en oxaloacetato mientras reduce NAD + a NADH. El oxaloacetato está entonces listo para combinarse con el siguiente acetil CoA para comenzar nuevamente el ciclo de Krebs (ver Figura\(\PageIndex{4}\)). Para cada giro del ciclo, se crean tres NADH, un ATP (a través de GTP) y un FADH 2. Cada carbono de piruvato se convierte en CO 2, el cual se libera como subproducto de la respiración oxidativa (aeróbica).
La fosforilación oxidativa y la cadena de transporte de electrones
La cadena de transporte de electrones (ETC) utiliza el NADH y el FADH 2 producidos por el ciclo de Krebs para generar ATP. Los electrones de NADH y FADH 2 se transfieren a través de complejos proteicos incrustados en la membrana mitocondrial interna mediante una serie de reacciones enzimáticas. La cadena de transporte de electrones consiste en una serie de cuatro complejos enzimáticos (Complejo I — Complejo IV) y dos coenzimas (ubiquinona y Citocromo c), que actúan como portadores de electrones y bombas de protones utilizadas para transferir iones H + al espacio entre las membranas mitocondriales interna y externa (Figura \(\PageIndex{5}\)). El ETC acopla la transferencia de electrones entre un donante (como NADH) y un aceptor de electrones (como O 2) con la transferencia de protones (iones H +) a través de la membrana mitocondrial interna, permitiendo el proceso de fosforilación oxidativa. En presencia de oxígeno, la energía se pasa, paso a paso, a través de los portadores de electrones para recoger gradualmente la energía necesaria para unir un fosfato al ADP y producir ATP. El papel del oxígeno molecular, O 2, es como aceptor de electrones terminal para la ETC. Esto significa que una vez que los electrones han pasado por todo el ETC, deben pasarse a otra molécula separada. Estos electrones, O 2 y H + iones de la matriz se combinan para formar nuevas moléculas de agua. Esta es la base de tu necesidad de respirar oxígeno. Sin oxígeno cesa el flujo de electrones a través del ETC.

Los electrones liberados de NADH y FADH 2 son pasados a lo largo de la cadena por cada uno de los portadores, los cuales se reducen cuando reciben el electrón y se oxidan al pasarlo al siguiente portador. Cada una de estas reacciones libera una pequeña cantidad de energía, que se utiliza para bombear iones H + a través de la membrana interna. La acumulación de estos protones en el espacio entre las membranas crea un gradiente protónico con respecto a la matriz mitocondrial.
También incrustado en la membrana mitocondrial interna hay un increíble complejo de poros proteicos llamado ATP sintasa. Efectivamente, es una turbina que es alimentada por el flujo de iones H + a través de la membrana interna hacia abajo en un gradiente y hacia la matriz mitocondrial. A medida que los iones H + atraviesan el complejo, el eje del complejo gira. Esta rotación permite que otras porciones de ATP sintasa alienten a ADP y P i a crear ATP. Al dar cuenta del número total de ATP producido por molécula de glucosa a través de la respiración aeróbica, es importante recordar los siguientes puntos:
- Una red de dos ATP se produce a través de la glucólisis (cuatro producidos y dos consumidos durante la etapa de consumo de energía). Sin embargo, estos dos ATP se utilizan para transportar el NADH producido durante la glucólisis desde el citoplasma hacia las mitocondrias. Por lo tanto, la producción neta de ATP durante la glucólisis es cero.
- En todas las fases posteriores a la glucólisis, el número de ATP, NADH y FADH 2 producidos debe multiplicarse por dos para reflejar cómo cada molécula de glucosa produce dos moléculas de piruvato.
- En la ETC, se producen alrededor de tres ATP por cada NADH oxidado. Sin embargo, solo se producen alrededor de dos ATP por cada FADH 2 oxidado. Los electrones de FADH 2 producen menos ATP, debido a que inician en un punto más bajo en el ETC (Complejo II) en comparación con los electrones del NADH (Complejo I) (ver Figura\(\PageIndex{5}\)).
Por lo tanto, por cada molécula de glucosa que ingresa a la respiración aeróbica, se produce un total neto de 36 ATPs (Figura\(\PageIndex{6}\)).

Gluconeogénesis
La gluconeogénesis es la síntesis de nuevas moléculas de glucosa a partir de piruvato, lactato, glicerol, o los aminoácidos alanina o glutamina. Este proceso se lleva a cabo principalmente en el hígado durante periodos de baja glucosa, es decir, bajo condiciones de ayuno, inanición y dietas bajas en carbohidratos. Entonces, ¿se puede plantear la pregunta de por qué el cuerpo crearía algo en lo que acaba de dedicarse una buena cantidad de esfuerzo a descomponer? Ciertos órganos clave, incluido el cerebro, pueden utilizar únicamente la glucosa como fuente de energía; por lo tanto, es esencial que el cuerpo mantenga una concentración mínima de glucosa en sangre. Cuando la concentración de glucosa en sangre cae por debajo de ese cierto punto, el hígado sintetiza nueva glucosa para elevar la concentración sanguínea a la normalidad.
La gluconeogénesis no es simplemente lo contrario de la glucólisis. Existen algunas diferencias importantes (Figura\(\PageIndex{7}\)). El piruvato es un material de partida común para la gluconeogénesis. Primero, el piruvato se convierte en oxaloacetato. El oxaloacetato sirve entonces como sustrato para la enzima fosfoenolpiruvato carboxiquinasa (PEPCK), que transforma el oxaloacetato en fosfoenolpiruvato (PEP). A partir de este paso, la gluconeogénesis es casi lo contrario de la glucólisis. PEP se convierte de nuevo en 2-fosfoglicerato, que se convierte en 3-fosfoglicerato. Luego, el 3-fosfoglicerato se convierte en 1,3 bisfosfosglicerato y luego en gliceraldehído-3-fosfato. Dos moléculas de gliceraldehído-3-fosfato se combinan para formar fructosa-1-6-bisfosfato, que se convierte en fructosa 6-fosfato y luego en glucosa-6-fosfato. Finalmente, una serie de reacciones genera la propia glucosa. En la gluconeogénesis (en comparación con la glucólisis), la enzima hexoquinasa es reemplazada por glucosa-6-fosfatasa, y la enzima fosfofructoquinasa-1 es reemplazada por fructosa-1,6-bisfosfatasa. Esto ayuda a la célula a regular la glucólisis y la gluconeogénesis independientemente entre sí.
Como se discutirá como parte de la lipólisis, las grasas se pueden descomponer en glicerol, el cual puede fosforilarse para formar fosfato de dihidroxiacetona o DHAP. El DHAP puede ingresar a la vía glucolítica o ser utilizado por el hígado como sustrato para la gluconeogénesis.

ENVEJECIMIENTO Y LA...
Tasa Metabólica del Cuerpo
La tasa metabólica del cuerpo humano disminuye casi 2 por ciento por década después de los 30 años. Los cambios en la composición corporal, incluida la reducción de la masa muscular magra, son los principales responsables de esta disminución. La pérdida más dramática de masa muscular, y consecuente disminución de la tasa metabólica, ocurre entre los 50 y 70 años de edad. La pérdida de masa muscular es el equivalente a una fuerza reducida, lo que tiende a inhibir a los adultos mayores de realizar suficiente actividad física. Esto da como resultado un sistema de retroalimentación positiva donde la actividad física reducida conduce a una mayor pérdida muscular, reduciendo aún más el metabolismo.
Hay varias cosas que se pueden hacer para ayudar a prevenir descensos generales en el metabolismo y para luchar contra la naturaleza cíclica de estos descensos. Estos incluyen desayunar, comer comidas pequeñas con frecuencia, consumir mucha proteína magra, beber agua para mantenerse hidratado, hacer ejercicio (incluido el entrenamiento de fuerza) y dormir lo suficiente. Estas medidas pueden ayudar a evitar que los niveles de energía bajen y frenar el impulso de aumentar el consumo de calorías por el exceso de bocadillos. Si bien estas estrategias no están garantizadas para mantener el metabolismo, sí ayudan a prevenir la pérdida muscular y pueden aumentar los niveles de energía. Algunos expertos también sugieren evitar el azúcar, lo que puede llevar al exceso de almacenamiento de grasa. Los alimentos picantes y el té verde también podrían ser beneficiosos. Debido a que el estrés activa la liberación de cortisol, y el cortisol ralentiza el metabolismo, evitar el estrés, o al menos practicar técnicas de relajación, también puede ayudar.
Revisión del Capítulo
Las enzimas metabólicas catalizan las reacciones catabólicas que descomponen los carbohidratos contenidos en los alimentos. La energía liberada se utiliza para alimentar las células y sistemas que componen tu cuerpo. El exceso o energía no utilizada se almacena como grasa o glucógeno para su uso posterior. El metabolismo de los carbohidratos comienza en la boca, donde la enzima amilasa salival comienza a descomponer los azúcares complejos en monosacáridos. Estos pueden ser transportados a través de la membrana intestinal hacia el torrente sanguíneo y luego a los tejidos corporales. En las células, la glucosa, un azúcar de seis carbonos, se procesa a través de una secuencia de reacciones en azúcares más pequeños, y se libera la energía almacenada dentro de la molécula. El primer paso del catabolismo de carbohidratos es la glucólisis, que produce piruvato, NADH y ATP. En condiciones anaerobias, el piruvato se puede convertir en lactato para mantener funcionando la glucólisis. En condiciones aeróbicas, el piruvato ingresa al ciclo de Krebs, también llamado ciclo del ácido cítrico o ciclo del ácido tricarboxílico. Además del ATP, el ciclo de Krebs produce moléculas FADH 2 y NADH de alta energía, que proporcionan electrones al proceso de fosforilación oxidativa que genera más moléculas de ATP de alta energía. Por cada molécula de glucosa que se procesa en la glucólisis, se puede crear una red de 36 ATP por respiración aeróbica.
En condiciones anaerobias, la producción de ATP se limita a las generadas por la glucólisis. Si bien se producen un total de cuatro ATP por glucólisis, se necesitan dos para comenzar la glucólisis, por lo que hay un rendimiento neto de dos moléculas de ATP.
En condiciones de baja glucosa, como ayuno, inanición o dietas bajas en carbohidratos, la glucosa se puede sintetizar a partir de lactato, piruvato, glicerol, alanina o glutamato. Este proceso, llamado gluconeogénesis, es casi el reverso de la glucólisis y sirve para crear moléculas de glucosa para órganos dependientes de glucosa, como el cerebro, cuando los niveles de glucosa caen por debajo de lo normal.
Preguntas de revisión
P. La glucólisis da como resultado la producción de dos moléculas ________ a partir de una sola molécula de glucosa. A falta de ________, el producto final de la glucólisis es ________.
A. acetil CoA, piruvato, lactato
B. ATP, carbono, piruvato
C. piruvato, oxígeno, lactato
D. piruvato, carbono, acetil CoA
Respuesta: C
P. El ciclo de Krebs convierte ________ a través de un ciclo de reacciones. En el proceso, se producen ATP, ________ y ________.
A. acetil CoA; FAD, NAD
B. acetil CoA; FADH 2; NADH
C. piruvato; NAD; FADH 2
D. piruvato; oxígeno; oxaloacetato
Respuesta: B
P. ¿Qué vía produce la mayor cantidad de moléculas de ATP?
A. Fermentación de ácido láctico
B. el ciclo de Krebs
C. la cadena de transporte de electrones
D. glucólisis
Respuesta: C
P. La respiración celular aeróbica resulta en la producción de estos dos productos.
A. NADH y FADH 2
B. ATP y piruvato
C. ATP y glucosa
D. ATP y H 2 O
Respuesta: D
P. Cuando NAD + se convierte en NADH, la coenzima ha sido ________.
A. reducido
B. oxidado
C. metabolizado
D. hidrolizado
Respuesta: A
Preguntas de Pensamiento Crítico
P. Explicar cómo se metaboliza la glucosa para producir ATP.
A. La glucosa se oxida durante la glucólisis, creando piruvato, el cual se procesa a través del ciclo de Krebs para producir NADH, FADH 2, ATP y CO 2. El FADH 2 y el NADH producen ATP.
P. La insulina se libera cuando se ingiere comida y estimula la captación de glucosa en la célula. Discutir el mecanismo que emplean las células para crear un gradiente de concentración que asegure la captación continua de glucosa del torrente sanguíneo.
A. Al ingresar a la célula, la hexoquinasa o la glucoquinasa fosforila la glucosa, convirtiéndola en glucosa-6-fosfato. De esta forma, la glucosa-6-fosfato queda atrapada en la célula. Debido a que toda la glucosa ha sido fosforilada, nuevas moléculas de glucosa pueden ser transportadas a la célula de acuerdo con su gradiente de concentración.
Glosario
- acetil coenzima A (acetil CoA)
- molécula de partida del ciclo de Krebs
- ATP sintasa
- complejo de poros proteicos que crea ATP
- respiración celular
- producción de ATP a partir de la oxidación de glucosa a través de la glucólisis, el ciclo de Krebs y la fosforilación oxidativa
- ciclo del ácido cítrico
- también llamado ciclo de Krebs o ciclo de ácido tricarboxílico; convierte el piruvato en CO 2 y moléculas de FADH 2, NADH y ATP de alta energía
- cadena de transporte de electrones (ETC)
- Vía de producción de ATP en la que los electrones pasan a través de una serie de reacciones de oxidación-reducción que forman agua y producen un gradiente de protones
- fase de consumo de energía
- primera fase de glucólisis, en la que son necesarias dos moléculas de ATP para iniciar la reacción
- fase de producción de energía
- segunda fase de la glucólisis, durante la cual se produce energía
- glucoquinasa
- enzima celular, que se encuentra en el hígado, que convierte la glucosa en glucosa-6-fosfato al ser absorbida en la célula
- gluconeogénesis
- proceso de síntesis de glucosa a partir de piruvato u otras moléculas
- glucosa-6-fosfato
- Glucosa fosforilada producida en la primera etapa de la glucólisis
- glucólisis
- serie de reacciones metabólicas que descomponen la glucosa en piruvato y producen ATP
- hexoquinasa
- enzima celular, que se encuentra en la mayoría de los tejidos, que convierte la glucosa en glucosa-6-fosfato al ser absorbida en la célula
- Ciclo de Krebs
- también llamado ciclo del ácido cítrico o ciclo del ácido tricarboxílico, convierte el piruvato en CO 2 y moléculas de FADH 2, NADH y ATP de alta energía
- monosacárido
- molécula de azúcar monomérica más pequeña
- fosforilación oxidativa
- proceso que convierte NADH y FADH 2 de alta energía en ATP
- polisacáridos
- carbohidratos complejos compuestos por muchos monosacáridos
- piruvato
- producto final de tres carbonos de glucólisis y material de partida que se convierte en acetil CoA que entra en el ciclo de Krebs
- amilasa salival
- enzima digestiva que se encuentra en la saliva y comienza la digestión de los carbohidratos en la boca
- aceptor de electrones terminal
- oxígeno, el receptor del hidrógeno libre al final de la cadena de transporte de electrones
Colaboradores y Atribuciones
- tricarboxylic acid cycle (TCA)
- also called the Krebs cycle or the citric acid cycle; converts pyruvate into CO2 and high-energy FADH2, NADH, and ATP molecules
- Template:ContribOpenStaxAP