
2.5: Acción de Neurotransmisores - Receptores Acoplados a Proteínas G
- Page ID
- 124526
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
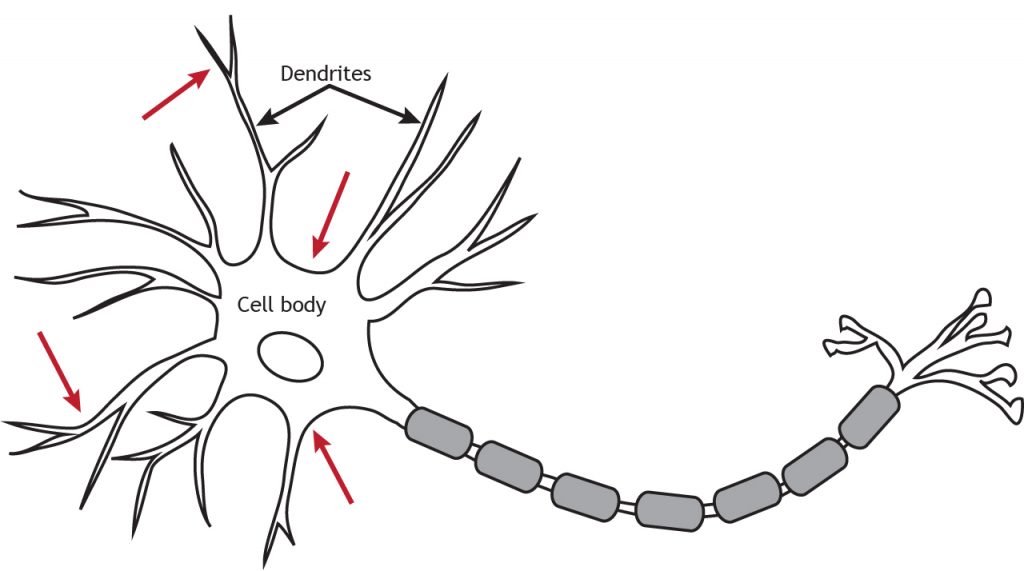
Los receptores acoplados a proteínas G (GPCR), también llamados receptores metabotrópicos, son proteínas unidas a la membrana que activan las proteínas G después de unirse a los neurotransmisores. Al igual que los receptores ionotrópicos, los receptores metabotrópicos se localizan principalmente a lo largo de las dendritas o el cuerpo celular, pero pueden estar presentes en cualquier lugar a lo largo de la neurona si hay una Los receptores metabotrópicos también son importantes para recibir información entrante de otras neuronas. Los GPCR tienen efectos más lentos que los receptores ionotrópicos, pero pueden tener efectos duraderos, a diferencia de la breve acción de un potencial postsináptico.
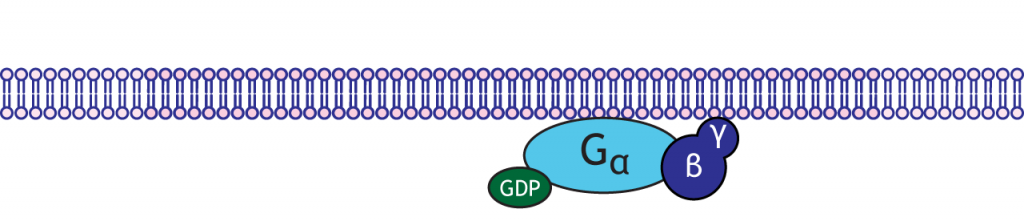
Proteínas G
Las proteínas G son enzimas con tres subunidades: alfa, beta y gamma. En el estado de reposo del complejo de proteína G, la subunidad alfa se une a una molécula GDP. Existen múltiples tipos de subunidades alfa, y cada una inicia diferentes cascadas celulares en la neurona.
Receptores acoplados a proteína G
Cuando un neurotransmisor se une a un GPCR, el receptor es capaz de interactuar con un complejo de proteína G inactivada. El complejo que se une es específico del receptor; diferentes receptores metabotrópicos para un mismo neurotransmisor pueden tener diferentes efectos en la célula debido a que se une la proteína G. Una vez acoplada al receptor, la molécula GDP se intercambia por una molécula GTP, y la proteína G se activa.
Animación 12.1. La unión del neurotransmisor a un receptor acoplado a proteína G provoca que el complejo de proteína G inactivada interactúe con el receptor. La molécula GDP se intercambia entonces por una molécula GTP, que activa el complejo de proteína G. 'G-Protein Binding' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 International License. Ver imagen estática de animación.
Después de la activación, el complejo de proteína G se separará en la subunidad alfa-GTP y la subunidad beta-gamma. Ambos componentes pueden alterar la función de las proteínas efectoras en la célula. Las funciones de las proteínas efectoras pueden variar desde alterar la permeabilidad iónica a través de la membrana abriendo canales iónicos hasta iniciar cascadas de segundos mensajeros. Las cascadas de segundos mensajeros pueden tener efectos celulares a largo plazo, generalizados y diversos, incluyendo la activación de enzimas celulares o la alteración de la transcripción génica.
Animación 12.2. Una vez activado, el complejo de proteína G se separará en la subunidad alfa-GTP y la subunidad beta-gamma. Estas subunidades pueden estimular o inhibir proteínas efectoras dentro de la célula. 'G-Protein Effects' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 International License. Ver imagen estática de animación.
Efectos celulares de las proteínas G
Canales Iones Abiertos — Subunidad Beta Gamma
En ciertas situaciones, la subunidad beta-gamma activada puede abrir o cerrar canales iónicos y cambiar la permeabilidad de la membrana. Los receptores muscarínicos de acetilcolina en el corazón utilizan esta vía. Cuando la acetilcolina se une a un receptor muscarínico en la fibra del músculo cardíaco, la subunidad beta-gamma activada abre un tipo de canal de potasio llamado canal de potasio acoplado a proteína G que rectifica hacia adentro (GIRK), hiperpolarizando la célula. Este efecto inhibitorio explica por qué la acetilcolina o un agonista como la atropina ralentizan la frecuencia cardíaca.
Animación 12.3. Algunos GPCR, como los receptores muscarínicos de acetilcolina en el corazón, alteran la permeabilidad celular al abrir canales iónicos. La subunidad beta-gamma activada del receptor muscarínico abre los canales de potasio GIRK y permite el eflujo de potasio. 'Canales Iónicos Beta-Gamma' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Segundo Mensajero Cascadas
Además de efectos directos como la subunidad beta-gamma activada abriendo canales iónicos, las proteínas G pueden tener muchas acciones indirectas en la célula mediante el uso de cascadas de segundos mensajeros. La vía específica del segundo mensajero que se activa o suprime por la acción de la proteína G depende del tipo de subunidad alfa.
Por ejemplo, la norepinefrina puede actuar sobre receptores alfa o beta-adrenérgicos. Los GPCR beta-adrenérgicos se acoplan a una proteína G estimuladora, o G s, que inicia el sistema de segundo mensajero AMP cíclico (AMPc) activando la enzima adenilciclasa. Los receptores alfa 2-adrenérgicos, sin embargo, se acoplan a una proteína G inhibitoria, o G i, y suprimen la actividad de la adenil ciclasa. Los receptores alfa 1-adrenérgicos se acoplan a un tercer tipo de proteína G, G q, que activa la vía de la fosfolipasa C. Un neurotransmisor puede, por lo tanto, causar una amplia gama de efectos celulares después de unirse a GPCR, a diferencia de la función única del flujo iónico a través de los receptores ionotrópicos. La vía iniciada por la norepinefrina dependerá del tipo de receptor que exprese una célula específica.
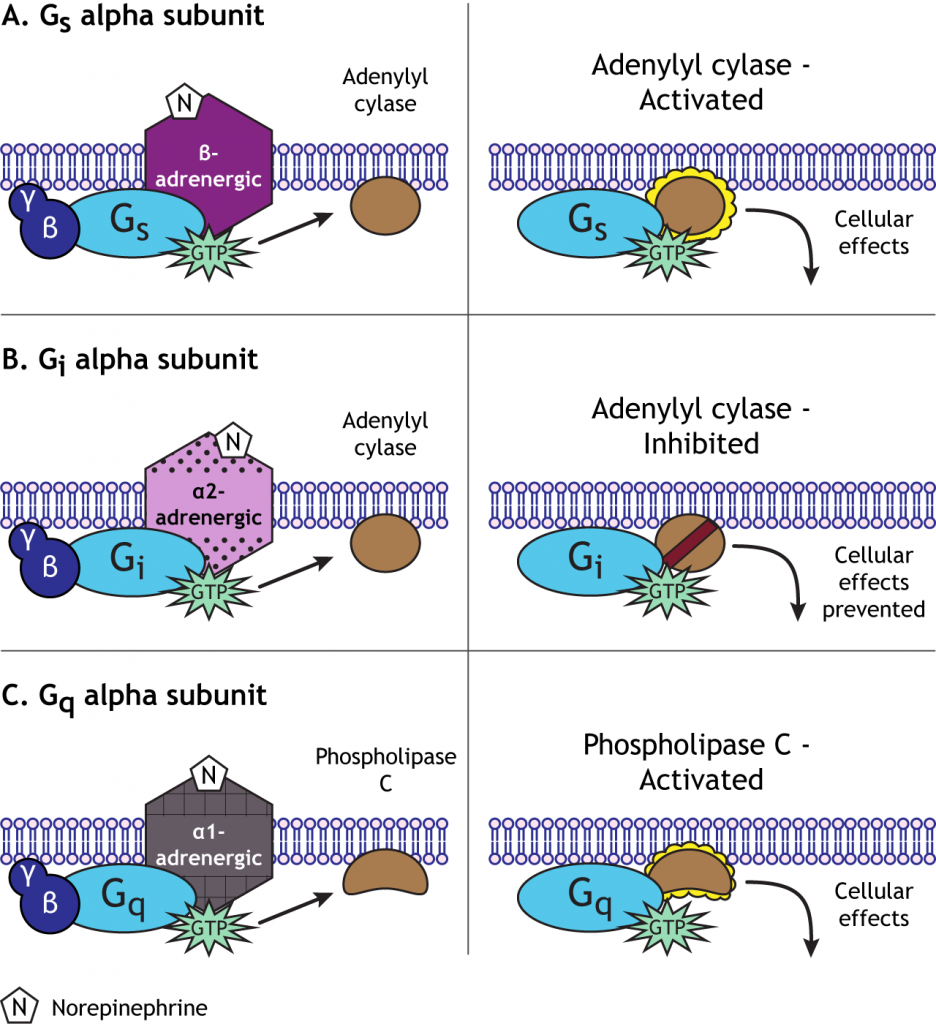
Cascada de Segundo Mensajero de AdenililCiclasa/
La ruta del segundo mensajero AMP cíclico (AMPc) es utilizada por muchos GPCR. La activación de la vía es causada por la subunidad alfa de G s y la inhibición de la vía es causada por la subunidad G i alfa. Cuando se activa, la adenilciclasa convierte el ATP en AMPc en el citoplasma, luego activa otra enzima llamada proteína quinasa A (PKA) uniéndose a las subunidades reguladoras, permitiendo que las subunidades catalíticas (funcionales) se separen y se activen. Las proteínas quinasas agregan una molécula de fosfato a las proteínas, un mecanismo llamado fosforilación. La adición del fosfato cambia la actividad de la proteína y cómo funciona en la célula.
Animación 12.4. Los GPCR que se acoplan a la subunidad alfa de G s inician la ruta adenilil ciclasa/AMPc. La subunidad G activa la adenil ciclasa, que luego convierte el ATP en AMPc. El cAMP se une y activa la proteína quinasa A (PKA), que fosforila las proteínas en la célula. 'Adenylyl Cyclasa Pathway' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Los efectos finales de esta vía dependerán de qué proteínas se dirigen. Por ejemplo, el AMPc puede abrir canales iónicos y PKA puede fosforilar canales iónicos alterando la permeabilidad y el potencial de membrana. La fosforilación puede abrir el canal, o puede modular la actividad del canal, haciendo que el canal sea más fácil de abrir o permanecer abierto por más tiempo.
Animación 12.5. La vía adenilil ciclasa/AMPc puede alterar muchas funciones celulares. Un ejemplo es que tanto AMPc como PKA pueden abrir canales iónicos. Al igual que los canales con puerta de enlace, también hay canales cerrados por campamento, que se abren después de la unión de cAMP. PKA es capaz de fosforilar y modular la función del canal iónico mediante la conversión de ATP en ADP. 'Second Messenger Ion Channel Action' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 International License. Ver imagen estática de animación.
Además de alterar la función del canal iónico, la PKA puede fosforilar otras proteínas importantes para la función de las neuronas, como las proteínas involucradas en la síntesis y liberación de neurotransmisores. Otro objetivo crítico de la fosforilación de PKA es el factor de transcripción CREB (proteína de unión al elemento de respuesta AMPc). Los factores de transcripción se unen al ADN en el núcleo y cambian la tasa de transcripción génica. La fosforilación por PKA puede hacer que CREB inicie la transcripción de genes, creando nuevas proteínas para la neurona. Dependiendo de qué genes se transcriban, los efectos sobre la neurona pueden ser duraderos.
En general, los neurotransmisores que trabajan a través de GPCR y cascadas de segundos mensajeros como la vía de la adenil ciclasa pueden causar una amplia gama de efectos celulares: desde la apertura de canales iónicos, hasta el cambio de la actividad proteica a través de la fosforilación, hasta la alteración de las proteínas sintetizadas en la neurona.
Animación 12.6. PKA puede fosforilar una serie de proteínas involucradas con la función de las neuronas. Puede dirigirse a proteínas involucradas con la síntesis, empaquetamiento y liberación de neurotransmisores, o puede ingresar al núcleo y fosforilar CREB, un factor de transcripción que puede iniciar la transcripción génica y la síntesis de proteínas. 'PKA Targets' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Fosfolipasa C/IP 3/DAG Segundo Mensajero Cascada
La subunidad alfa de Gq inicia una vía de señalización separada en la célula activando la fosfolipasa C. La fosfolipasa C se dirige a PIP 2 (fosfatidilinositol 4,5-bisfosfato), que es un fosfolípido presente en la membrana plasmática de la célula. El PIP 2 se divide en dos moléculas celulares: IP 3 (inositol 1,4,5-trisfosfato) y DAG (diacilglicerol). DAG permanece en la membrana e interactúa con la proteína quinasa c (PKC). IP 3 se mueve al retículo endoplásmico donde abre los canales de calcio y permite que el calcio fluya hacia el citosol.
El calcio también es un segundo mensajero en la célula. Un efecto importante es la unión del calcio a la proteína calmodulina. Este complejo puede entonces activar otra quinasa, la proteína quinasa dependiente de calcio/calmodulina (CaMK). Tanto PKC como CaMK pueden fosforilar proteínas celulares y nucleares específicas como PKA.
Animación 12.7. La subunidad de proteína G q activa la fosfolipasa C, que convierte el fosfolípido PIP 2 en la membrana celular en DAG, otra molécula unida a la membrana, e IP 3, una molécula citoplásmica. DAG puede interactuar con PKA, iniciando la fosforilación de proteínas celulares. IP 3 abre canales de calcio en el retículo endoplásmico, permitiendo que el calcio fluya hacia el citoplasma. El calcio, otro segundo mensajero puede tener muchos efectos celulares. Puede unirse a la calmodulina, que luego activa CaMK, provocando la fosforilación de más dianas proteicas. 'IP 3-DAG Pathway' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
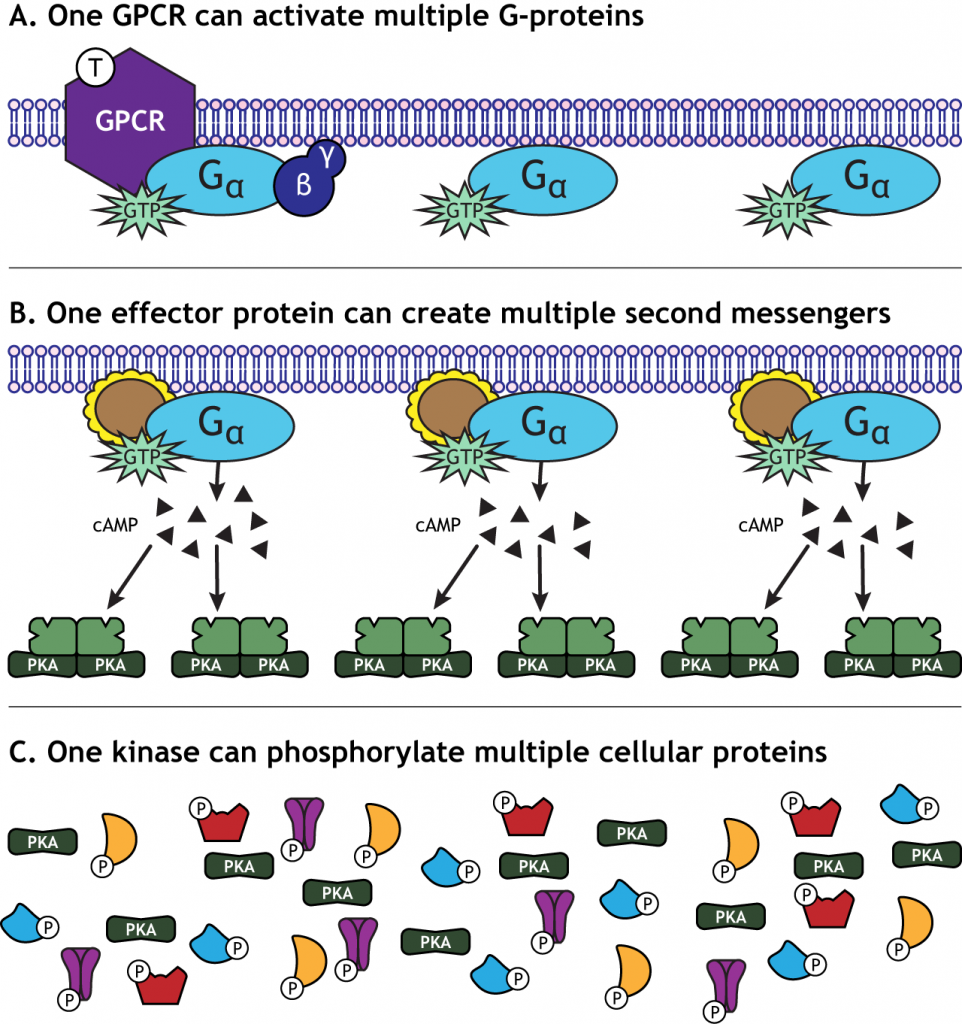
Amplificación de señal
Una característica de la activación del GPCR es la amplificación de la señal que tiene lugar. Un receptor es capaz de activar más de un complejo de proteína G. La proteína efectora activada por la proteína G puede crear muchos segundos mensajeros, y las proteínas quinasas activadas pueden fosforilar múltiples proteínas celulares. Esto significa que un neurotransmisor puede tener un efecto significativo en la función celular.
Terminación de señal
Finalmente, la cascada iniciada por la unión del neurotransmisor al GPCR necesita terminar. La subunidad alfa de la proteína G es capaz de convertir el GTP unido de nuevo a GDP después de un corto período de tiempo, inactivando la proteína G. La subunidad alfa interactuará entonces con una subunidad beta-gamma y permanecerá en estado de reposo hasta ser activada por otro GPCR. Las enzimas en la célula llamadas proteínas fosfatasas encuentran y eliminan los grupos fosfato agregados a las proteínas celulares por las proteínas quinasas. Y finalmente, existen otros mecanismos celulares para eliminar el calcio del citoplasma y degradar a otros segundos mensajeros.
Claves para llevar
- Los receptores acoplados a proteínas G se basan en la activación de las proteínas G para causar cambios celulares
- Los receptores acoplados a proteína G tienen efectos más lentos que los receptores activados por ligando
- Las proteínas G pueden abrir canales iónicos, alterar la función de la proteína a través de la fosforilación y alterar la transcripción génica
- La subunidad G inicia la vía de señalización de adenilil ciclasa/AMPc
- La subunidad G i inhibe la vía de señalización de adenilil ciclasa/AMPc
- La subunidad G q inicia la vía de señalización de fosfolipasa C/IP 3/DAG
¡Ponte a prueba!
Un elemento H5P interactivo ha sido excluido de esta versión del texto. Puedes verlo en línea aquí:
https://openbooks.lib.msu.edu/neuroscience/?p=397#h5p-13
Revisión adicional
¿Cuáles son algunas diferencias entre los receptores neurotransmisores ionotrópicos y metabotrópicos?
RESPUESTAS
Versión en video de la lección

Un elemento de YouTube ha sido excluido de esta versión del texto. Puedes verlo en línea aquí: https://openbooks.lib.msu.edu/neuroscience/?p=397