
2.4: Acción de Neurotransmisores - Receptores Ionotrópicos
- Page ID
- 124536
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Los receptores ionotrópicos, también llamados canales activados por neurotransmisores o activados por ligandos, son canales iónicos que se abren en respuesta a la unión de un neurotransmisor. Se localizan principalmente a lo largo de las dendritas o cuerpo celular, pero pueden estar presentes en cualquier lugar a lo largo de la neurona si hay una sinapsis. Los canales controlados por ligando son importantes para recibir información entrante de otras neuronas.
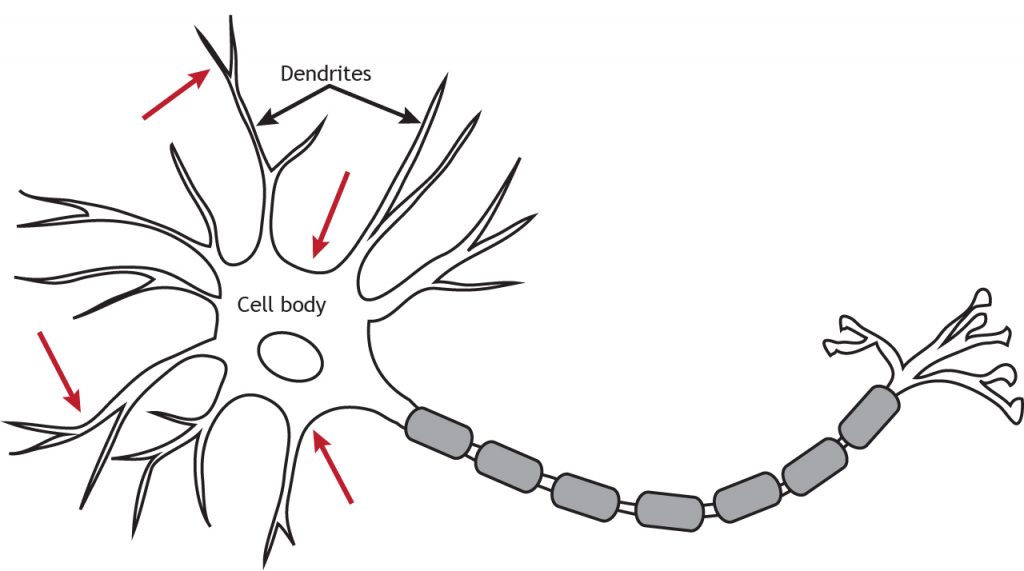
Aunque los receptores ionotrópicos son canales iónicos, se abren de una manera diferente a los canales iónicos regulados por voltaje necesarios para la propagación del potencial de acción. Los receptores ionotrópicos están regulados por ligando, lo que significa que una molécula específica, como un neurotransmisor, debe unirse al receptor para hacer que el canal se abra y permita el flujo de iones. Como se vio en capítulos anteriores, los canales regulados por voltaje se abren en respuesta a que el potencial de membrana alcanza el umbral.
Animación 11.1. Los receptores ionotrópicos, también llamados canales regulados por ligandos, son canales iónicos que se abren por la unión de neurotransmisores. Los canales regulados por voltaje se abren por el potencial de membrana de la celda que alcanza el umbral. Ambos tipos de canales permiten que los iones se difundan por su gradiente electroquímico. Los canales forrados, verde azulado representan receptores de glutamato; los canales amarillos sólidos representan receptores GABA; los canales punteados y azules representan canales de sodio regulados por voltaje. 'Ion Channel Gating' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Los receptores solo pueden ser abiertos por un ligando específico. Los neurotransmisores y receptores encajan como una cerradura y llave; solo ciertos neurotransmisores son capaces de unirse y abrir ciertos receptores.
Animación 11.2. Dado que los receptores de neurotransmisores solo pueden unirse a neurotransmisores específicos, el glutamato se une y abre los receptores de glutamato pero no tiene ningún efecto sobre los receptores GABA. Los canales forrados, verde azulado representan receptores de glutamato; los canales amarillos sólidos representan receptores GABA. 'Ligando y Receptor' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Receptores de glutamato
El glutamato es el neurotransmisor excitador primario en el sistema nervioso central y abre canales catiónicos no selectivos. Existen tres subtipos de receptores de glutamato. Los receptores AMPA (ácido α-amino-3-hidroxi-5-metil-4-isoxazolpropiónico) y kainato permiten que tanto el sodio como el potasio crucen la membrana. Aunque el potasio puede salir de la célula cuando los receptores se abren, el gradiente electroquímico que impulsa el movimiento del ion sodio es más fuerte que el gradiente que impulsa el movimiento del potasio, lo que resulta en una despolarización del potencial de la membrana.
Animación 11.3. Los receptores AMPA y kainato glutamato son canales iónicos no selectivos que permiten que tanto el sodio como el potasio fluyan a través de la membrana. Cuando el glutamato se une, el sodio entra y el potasio fluye hacia afuera. El canal verde azulado alineado representa receptores SamPA; el canal verde azulado a cuadros representa receptores de kainato. 'AMPA y Kainate' de Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
El receptor NMDA (N-metil-D-aspartato) requiere la unión del glutamato para abrirse, pero también depende del voltaje. Cuando el potencial de membrana está por debajo, en o cerca de reposo, un ion magnesio bloquea el receptor abierto de NMDA y evita que otros iones se muevan a través del canal. Una vez que la célula se despolariza, el bloque de magnesio es expulsado del receptor, lo que permite que el sodio, el potasio y el calcio crucen la membrana. El cambio de voltaje necesario para abrir el receptor NMDA suele ser el resultado de la activación del receptor AMPA. El glutamato liberado se une tanto a los receptores AMPA como a NMDA, la afluencia de sodio se produce a través de canales AMPA abiertos, lo que despolariza la célula lo suficiente como para expulsar el ion magnesio y permitir el flujo de iones a través de los receptores NMDA.
Animación 11.4. Los receptores NMDA se abren mediante una combinación de unión de glutamato y un disparador de voltaje. A niveles bajos de estimulación, cuando el potencial de la membrana está cerca de reposo, un ion magnesio bloquea el canal abierto del receptor NMDA impidiendo el flujo iónico. Los iones pueden fluir a través de receptores AMPA abiertos, los cuales comienzan a despolarizar la membrana. El cambio de voltaje eventualmente expulsa el ion magnesio del canal, permitiendo que el sodio, el potasio y el calcio crucen la membrana. El canal verde azulado alineado representa los receptores AMPA; el canal violeta punteado representa los receptores NMDA. 'AMPA y NMDA' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Receptores nicotínicos de acetilcolina
Como los receptores de glutamato, los receptores nicotínicos de acetilcolina son canales catiónicos no selectivos. Los receptores nicotínicos, sin embargo, se localizan principalmente fuera del sistema nervioso central. Los receptores nicotínicos se utilizan en la unión neuromuscular
Receptores GABA y Glicina
Los receptores GABA y glicina son canales de cloruro. Dado que un aumento de la permeabilidad al cloruro a través de la membrana es inhibitorio, la unión de GABA o glicina a su receptor ionotrópico respectivo provocará inhibición.
Animación 11.5. GABA y glicina son receptores inhibidores que son selectivos al cloruro. El canal amarillo sólido representa un receptor GABA; el canal amarillo modelado representa un receptor de glicina. 'GABA y Glicina' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Los receptores ionotrópicos causan potenciales postsinápticos
Los potenciales postsinápticos (Capítulo 5) son el resultado de la apertura de receptores ionotrópicos. Los receptores ionotrópicos excitadores aumentan la permeabilidad al sodio a través de la membrana, mientras que los receptores ionotrópicos inhibidores incrementan El flujo iónico a través de los receptores ionotrópicos sigue los mismos principios que otros canales iónicos cubiertos hasta ahora.
Revisión del potencial de equilibrio
Anteriormente, cubrimos el movimiento de iones a través de canales regulados por voltaje y discutimos que los gradientes electroquímicos conducirán el movimiento iónico hacia el equilibrio. El potencial de membrana de la neurona en el que se produce el equilibrio y equilibrio de los gradientes químicos y eléctricos es el potencial de equilibrio del ión.
Animación 11.6. Los iones se mueven a través de canales abiertos de voltaje tratando de alcanzar el equilibrio. A medida que los iones atraviesan la membrana, el potencial de membrana de la neurona se acerca al potencial de equilibrio del ion. En la animación, se abre un canal de sodio regulado por voltaje y el sodio fluye hasta que el potencial de membrana equivale aproximadamente a +60 mV, potencial de equilibrio del sodio. El canal punteado azul representa un canal de sodio dependiente de voltaje. 'Potencial de Equilibrio' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Potencial de inversión
Este mismo principio se utiliza para el movimiento iónico a través de receptores ionotrópicos. El potencial de membrana en el que el flujo iónico a través de un receptor está en equilibrio se denomina potencial de inversión del receptor. La dirección del movimiento iónico se puede predecir si se conoce el potencial de inversión del receptor.
GABA y Glicina — Receptores Selectivos a Un Ion
Cuando se abre un receptor ionotrópico que es selectivo a un solo ion, el potencial de inversión del receptor es el mismo que el potencial de equilibrio del ion. Los receptores GABA y glicina solo permiten que los iones cloruro crucen la membrana. Por lo tanto, el potencial de inversión de un receptor GABA o glicina es igual al potencial de equilibrio del cloruro, y la unión de GABA o glicina a su respectivo receptor ionotrópico provocará un potencial postsináptico inhibitorio (IPSP).
Animación 11.7. Los iones se mueven a través de canales abiertos con ligandos tratando de alcanzar el equilibrio. A medida que los iones atraviesan la membrana, el potencial de membrana de la neurona se acerca al potencial de inversión del receptor. Cuando el receptor ionotrópico solo aumenta la permeabilidad para un ion, el potencial de inversión del receptor es el mismo que el potencial de equilibrio del ión. En la animación, un receptor GABA se abre y el cloruro fluye hasta que el potencial de membrana es igual a aproximadamente -65 mV, potencial de inversión de GABA y potencial de equilibrio de cloruro. El aumento de la permeabilidad al cloruro provoca un IPSP e inhibe la neurona. El canal amarillo a cuadros representa un receptor GABA. 'Potencial de inversión GABA' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Glutamato - Potencial de inversión para receptores que no son selectivos
Sin embargo, si el receptor ionotrópico permite el flujo de más de un ion, o no es selectivo, el potencial de inversión del receptor no es igual al potencial de equilibrio de ninguno de los iones sino que está en algún punto intermedio. El potencial de equilibrio del sodio es aproximadamente +60 mV, y el potencial de equilibrio del potasio es aproximadamente -80 mV. Un receptor de glutamato es un canal catiónico no selectivo que permite el flujo de ambos iones, y el potencial de inversión del receptor es de 0 mV. Esto significa que si el potencial de membrana de la neurona es negativo, las fuerzas impulsoras que actúan sobre el sodio son más fuertes que las fuerzas impulsoras que actúan sobre el potasio, por lo que fluirá más sodio que el potasio fluirá hacia afuera, y el potencial de membrana se despolarizará, provocando un potencial postsináptico excitatorio (EPSP).
Animación 11.8. El potencial de inversión de un receptor ionotrópico que no es selectivo a un ion caerá entre los potenciales de equilibrio de los iones permeables. Los receptores de glutamato permiten el flujo de iones tanto de sodio como de potasio, por lo que el potencial de inversión para el receptor es de aproximadamente 0 mV. Más sodio fluirá hacia la célula que el potasio fluye hacia afuera, resultando en una despolarización de la membrana. La línea, canal verde azulado representa un receptor de glutamato. 'Potencial de inversión de glutamato — Descanso' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Si el potencial de membrana alcanzara el potencial de inversión del receptor de glutamato, los gradientes electroquímicos que actúan sobre el sodio y el potasio se equilibrarían, por lo que el flujo global de iones en ambas direcciones sería igual, y el potencial de membrana no cambiaría.
Animación 11.9. En el potencial de inversión, no hay flujo iónico neto en ninguna dirección. Un número igual de iones de sodio ingresa a la célula a medida que los iones de potasio salen. Dado que no hay cambio en el voltaje en el potencial de inversión, si el receptor permanece abierto, el potencial de membrana permanecería en 0 mV. 'Potencial de inversión de glutamato — 0 mV' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC-BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Claves para llevar
- Los receptores ionotrópicos son canales iónicos regulados por ligando que se abren cuando se une un neurotransmisor específico
- Para receptores selectivos a un ion, el potencial de inversión es igual al potencial de equilibrio del ión
- Para receptores no selectivos para un solo ion, el potencial de inversión es un valor entre los potenciales de equilibrio de los iones
- El glutamato es un neurotransmisor excitatorio que abre canales catiónicos no selectivos que permiten la afluencia de sodio, provocando una EPSP
- GABA y glicina son neurotransmisores inhibidores que abren canales de cloruro, provocando una IPSP
¡Ponte a prueba!
Un elemento H5P interactivo ha sido excluido de esta versión del texto. Puedes verlo en línea aquí:
https://openbooks.lib.msu.edu/neuroscience/?p=368#h5p-12
Revisión adicional
Una neurona postsináptica (Célula A) está en reposo a -60 mV y recibe entrada de cinco neuronas glutamato separadas y una neurona GABA. Los cambios en el potencial de membrana postsináptica se pueden medir mediante un electrodo de registro localizado en el cuerpo celular.
Dibuja el cambio en el potencial de membrana postsináptica que esperarías ver después de cada una de las siguientes manipulaciones:
- Una neurona de glutamato presináptica dispara un potencial de acción y libera neurotransmisor
- La neurona GABA presináptica dispara un potencial de acción y libera neurotransmisor
- Una neurona de glutamato presináptica dispara cinco potenciales de acción y libera neurotransmisor
RESPUESTAS
Versión en video de la lección

Un elemento de YouTube ha sido excluido de esta versión del texto. Puedes verlo en línea aquí: https://openbooks.lib.msu.edu/neuroscience/?p=368