2.7: Ácidos grasos
- Page ID
- 53134

A diferencia de los monosacáridos, nucleótidos y aminoácidos, los ácidos grasos no son monómeros que se unen entre sí para formar moléculas mucho más grandes. Aunque los ácidos grasos se pueden unir entre sí, por ejemplo, en triacilgliceroles o fosfolípidos, no están unidos directamente entre sí, y generalmente no más de tres en una molécula dada. Los ácidos grasos en sí son largas cadenas de átomos de carbono rematadas con un grupo carboxilo. La longitud de la cadena puede variar, aunque la mayoría están entre 14 y 20 carbonos, y en orden superior plantas y animales, los ácidos grasos con 16 y 18 carbonos son las especies principales.
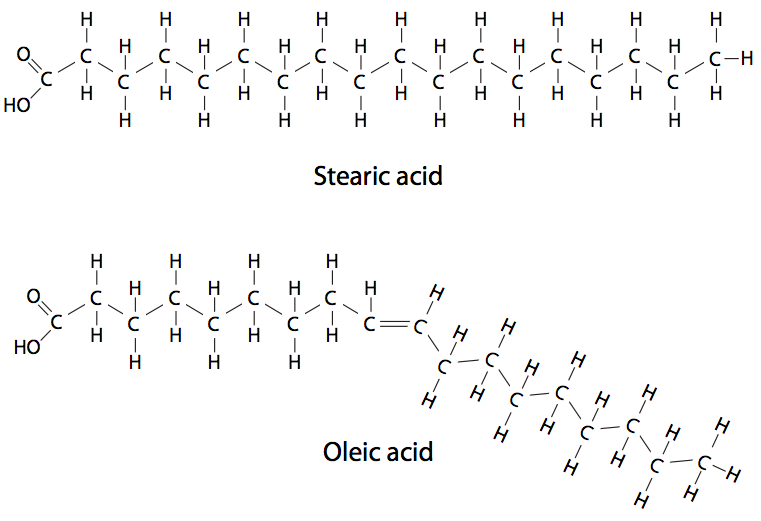
Debido al mecanismo de síntesis, la mayoría de los ácidos grasos tienen un número par de carbonos, aunque también se pueden generar cadenas de carbono impares. Se puede generar más variedad por dobles enlaces entre los carbonos. Las cadenas de ácidos grasos sin dobles enlaces están saturadas, porque cada carbono está saturado con tantos átomos de hidrógeno unidos como sea posible. Las cadenas de ácidos grasos con dobles enlaces son insaturadas (Figura\(\PageIndex{13}\)). Aquellos con más de un doble enlace se denominan poliinsaturados. Los ácidos grasos en las células eucariotas se dividen casi uniformemente entre los tipos saturados e insaturados, y muchos de estos últimos pueden ser poliinsaturados. En procariotas, la poliinsaturación es rara, pero otras modificaciones como la ramificación y la ciclación son más comunes que en los eucariotas. A continuación se muestra una tabla de ácidos grasos comunes.
| Ácido mirístico | 14:0 (14 carbonos, sin dobles enlaces |
| Ácido palmítico | 16:0 |
| Ácido esteárico | 18:0 |
| Ácido araquídico | 20:0 |
| Ácido Palmitoleico | 16:1 |
| Ácido oleico | 18:1 |
| Ácido linoleico | 18:2 |
| Ácido araquidónico | 2:4 |
Existen diferencias físicas significativas entre los ácidos grasos saturados e insaturados debido simplemente a la geometría de los carbonos de doble enlace. Un ácido graso saturado es muy flexible con rotación libre alrededor de todos sus enlaces C-C. Los diagramas lineales y fórmulas habituales que representan ácidos grasos saturados también sirven para explicar la capacidad de los ácidos grasos saturados para empaquetarse estrechamente, con muy poco espacio intermedio. Los ácidos grasos insaturados, por otro lado, no pueden empaquetarse con tanta fuerza debido a la restricción rotacional impoartada por el doble enlace. Los carbonos no pueden rotar alrededor del doble enlace, por lo que ahora hay una “torcedura” en la cadena. Generalmente, los carbonos de doble enlace en los ácidos grasos están en la configuración cis, introduciendo una curvatura de 30 grados en la estructura.
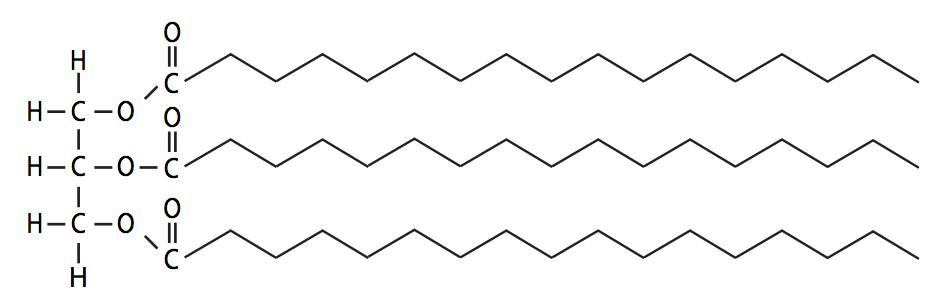
Los ácidos grasos dentro de las células suelen ser partes de moléculas más grandes, en lugar de ácidos libres. Algunos de los lípidos más comunes derivados de los ácidos grasos son triacilgliceroles, fosfoglicéridos y esfingolípidos. Los triacilgliceroles, como su nombre lo indica, son tres cadenas de ácidos grasos (acilo) conectadas a una molécula de glicerol por enlaces éster (Figura\(\PageIndex{14}\)). Los triacilgliceroles, también conocidos como triglicéridos, pueden tener ácidos grasos de los mismos (triacilgliceroles simples) o de diferentes tipos (triacilgliceroles mixtos). Las mezclas de estas son las principales moléculas de almacenamiento de energía a largo plazo para la mayoría de los Si bien pueden denominarse coloquialmente grasas o aceites, la única diferencia real es el grado de saturación de sus ácidos grasos constituyentes. Las mezclas con mayores porcentajes de ácidos grasos saturados tienen un punto de fusión más alto y si son sólidas a temperatura ambiente, se les denomina grasas. Las mezclas de triacilglicerol que quedan líquidas a temperatura ambiente son aceites.
En medicina humana, una prueba común para los factores de riesgo de enfermedades cardíacas es la medición de los niveles de triglicéridos en la sangre. Si bien varios tipos de células pueden producir y usar triglicéridos, la mayoría de los triglicéridos en las personas se concentran en el tejido adiposo, que está compuesto por adipocitos, o células grasas, aunque el hígado también es un importante almacén de grasa. Estas células se han especializado para portar glóbulos de grasa que ocupan la mayor parte del volumen de la célula. Cuando los niveles de triglicéridos en la sangre son altos, significa que la grasa se está produciendo o ingeriendo más rápido de lo que pueden ser absorbidos por los adipocitos.
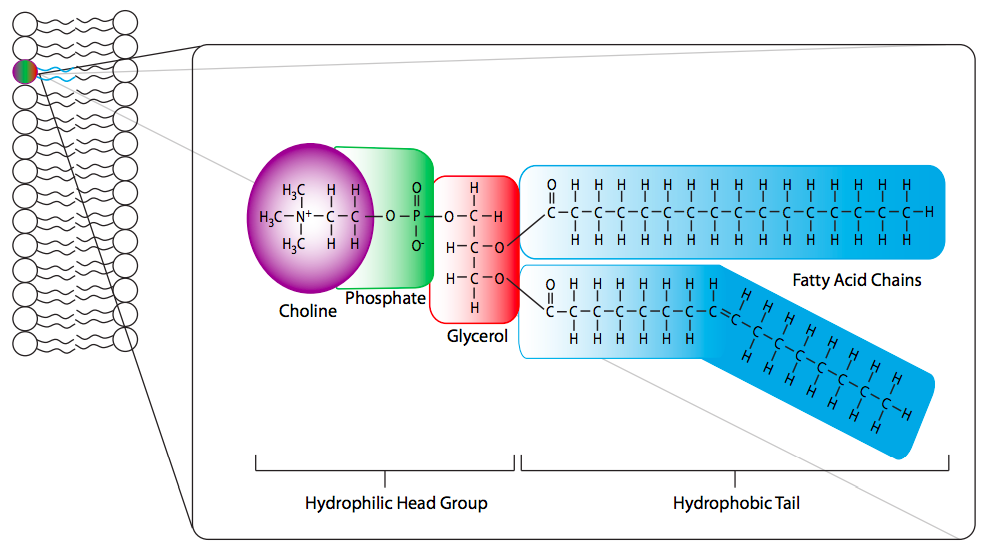
Los fosfolípidos (también llamados fosfoglicéridos o glicerofosfolípidos), también se basan en la unión de ácidos grasos al glicerol. Sin embargo, en lugar de tres colas de acilo graso, solo hay dos, y en la tercera posición se encuentra un grupo fosfato (Figura\(\PageIndex{15}\)). El grupo fosfato también se une a un “grupo principal”. La identidad del grupo de cabeza nombra a la molécula, junto con las colas de acilo graso. En la Figura de ejemplo, 1-estearoílo se refiere al ácido esteárico en el 1-carbono de la cadena principal de glicerol; 2-palmitoílo se refiere al ácido palmítico en el 2-carbono del glicerol, y fosfatidiletanolamina se refiere al grupo fosfato y su etanolamina unida, que están unidos al glicerol 3-carbono. Debido al grupo fosfato de carga negativa y a un grupo de cabeza que a menudo es polar o cargado, los fosfolípidos son anfipáticos, portando un fuerte carácter hidrófobo en las dos colas de acilo graso y un fuerte carácter hidrófilo en el grupo de cabeza. Esta anfipaticidad es crucial en el papel de los fosfolípidos como componente principal de las membranas celulares.
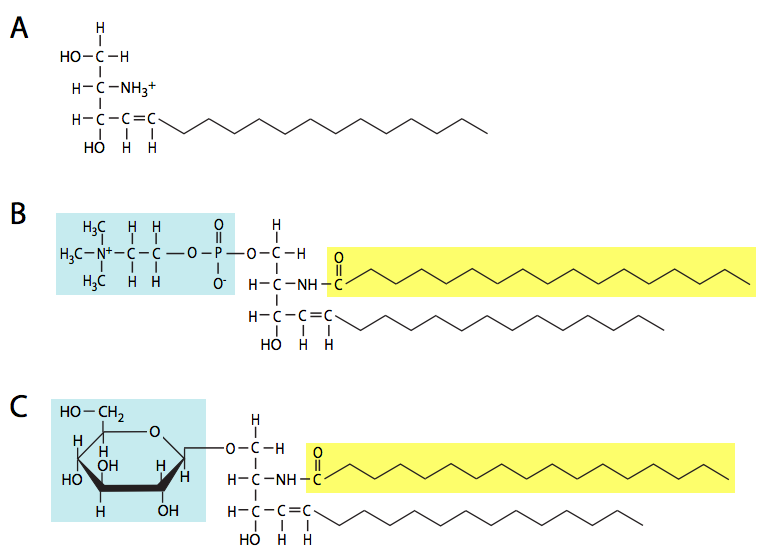
Los esfingolípidos (Figura\(\PageIndex{16}\)) también son constituyentes importantes de las membranas, y se basan no en una cadena principal de glicerol, sino en el aminoalcohol, esfingosina (o dihidroesfingosina). Hay cuatro tipos principales de esfingolípidos: ceramidas, esfingomielinas, cerebrosidos y gangliósidos. Las ceramidas son moléculas de esfingosina con una cola de ácido graso unida al grupo amino. Las esfingomielinas son ceramidas en las que se une una fosfocolina o fosfoetanolamina al carbono 1. Los cerebrosidos y gangliósidos son glicolípidos; tienen un azúcar o azúcares, respectivamente, unidos al 1-carbono de una ceramida. Todos los oligosacáridos unidos a gangliósidos contienen al menos un residuo de ácido siálico. Además de ser un componente estructural de la membrana celular, los gangliósidos son particularmente importantes en el reconocimiento de célula a célula.
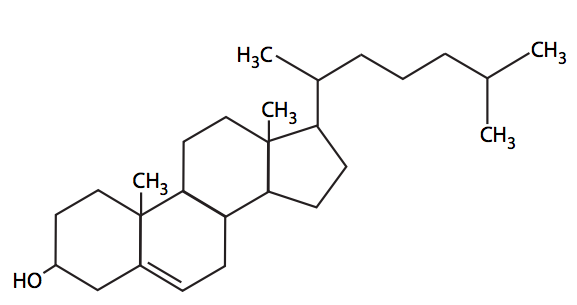
Los lípidos se definen vagamente como compuestos biológicos que son insolubles en agua pero que son solubles en disolventes orgánicos como el metanol o el cloroformo. Esto incluye los derivados de ácidos grasos enumerados anteriormente, e incluye el tema final para este capítulo, el colesterol. El colesterol (Figura\(\PageIndex{17}\)) es el principal derivado biológico del ciclopentanoperhidrofenantreno, un hidrocarburo saturado que consta de cuatro formaciones de anillos fusionados. Es un componente importante de las membranas plasmáticas en las células animales, y también es el precursor metabólico de las hormonas esteroides, como el cortisol o el b-estradiol. Las células vegetales tienen poco o ningún colesterol, pero otros esteroles como el estigmasterol están presentes. De igual manera, los hongos tienen sus esteroles particulares. Sin embargo, los procariotas no contienen, en su mayor parte, ninguna molécula de esterol.