
12.4: Genes supresores de tumores
- Page ID
- 56574

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Algunos genes suprimen la formación de tumores.
- Su producto proteico inhibe la mitosis.
- Cuando muta, el alelo mutante se comporta como un recesivo; es decir, mientras la célula contenga un alelo normal, la supresión tumoral continúa. (Los oncogenes, por el contrario, se comportan como dominantes; un alelo mutante, o demasiado activo, puede predisponer la célula a la formación de tumores).
RB - el gen del retinoblastoma
El retinoblastoma es un tumor canceroso de la retina. Ocurre en dos formas:
- Retinoblastoma familiar: Tumores múltiples en las retinas de ambos ojos que ocurren en las primeras semanas de la infancia.
- Retinoblastoma esporádico: Un solo tumor aparece en un ojo en algún momento de la primera infancia antes de que la retina esté completamente desarrollada y cese la mitosis en ella.
Retinoblastoma familiar
El retinoblastoma familiar ocurre cuando un bebé hereda de uno de sus padres un cromosoma (número 13) que tiene su locus RB eliminado (o mutado de otra manera). La proteína Rb normal controla el ciclo celular. Integra las señales que llegan a la célula para determinar si es seguro para la célula completar el paso de G 1 del ciclo celular a la mitosis.
Mecanismo
La proteína Rb no fosforilada impide que las células entren en la fase S del ciclo celular. Lo hace uniéndose a factores de transcripción llamados E2F. Esto evita que las E2Fs se unan a los promotores de proto-oncogenes tales como c- myc y c- fos. La transcripción de c- myc y c- fos es necesaria para la mitosis, por lo que bloquear los factores de transcripción necesarios para activar estos genes evita la división celular. Sin embargo, si las condiciones son adecuadas para que la célula complete con éxito la mitosis, la proteína Rb se fosforila, libera las E2Fs y la célula puede proceder a través del ciclo celular.
La proteína Rb también juega un papel en la mitosis misma: es necesaria para la condensación cromosómica adecuada a partir de la profase, así como su correcta unión al huso. La falla de la función de Rb durante la mitosis puede provocar aneuploidía y rotura cromosómica.

Una mutación aleatoria del locus RB restante en cualquier célula retiniana —que son células que no se dividen y no deben entrar en el ciclo celular— elimina completamente la inhibición proporcionada por la proteína Rb, y la célula afectada se convierte en un tumor. Entonces, en esta forma de la enfermedad, una mutación de la línea germinal más una mutación somática del segundo alelo conduce a la enfermedad.
Retinoblastoma esporádico
En esta enfermedad, ambos genes RB heredados son normales y una sola célula debe ser tan desafortunada como para sufrir una mutación somática (a menudo una deleción) en ambos para convertirse en un tumor. Tal doble golpe es un evento extremadamente improbable, y por lo tanto solo raramente ocurrirá tal tumor. (En ambas formas de la enfermedad, se puede salvar la vida del paciente si el tumor (es) se detecta (n) pronto y se extirpa el (los) ojo (s) afectado (s).)
p53
El producto del gen supresor tumoral p53 es una proteína de 53 kilodaltons (de ahí el nombre). (Encontrará que el gen humano se designa de diversas maneras como P53, TP53 ["proteína tumoral 53"] y TRP53 [" proteína relacionada con la transformación 53"])
La proteína p53 evita que una célula complete el ciclo celular si su ADN está dañado o la célula ha sufrido otro tipo de daño.
Cuando
- el daño es menor, p53 detiene el ciclo celular —de ahí la división celular— hasta que se repare el daño.
- el daño es mayor y no se puede reparar, p53 desencadena que la célula se suicide por apoptosis.
Estas funciones hacen de p53 un actor clave para protegernos contra el cáncer; es decir, es un importante gen supresor de tumores. Más de la mitad de todos los cánceres humanos de pulmón, ovario y colorrectal albergan mutaciones p53 y no tienen proteína p53 funcional.
Los ratones han sido curados del cáncer tratándolos con un péptido que activa la producción de la proteína p53 en las células tumorales. Sin embargo, puede haber una compensación involucrada: el exceso de producción de la proteína p53 conduce a un envejecimiento acelerado en ratones.
Los elefantes son muy longevos pero rara vez desarrollan cánceres. Resulta que sus células contienen 40 copias del gen p53 (TP53) en comparación con las dos que tenemos nosotros y otros mamíferos.
p16 Ink4a
El producto del gen supresor tumoral INK4a es una proteína de 16 kilodaltons (de ahí el nombre).
Al igual que p53, bloquea la progresión a través del ciclo celular —en este caso inhibiendo la acción de la quinasa dependiente de ciclina Cdk4.
A medida que un animal envejece, sus células producen cantidades crecientes de p16 Ink4a. Esto es probablemente algo bueno ya que reduce el riesgo de que la célula entre en mitosis incontrolada, es decir, convertirse en cáncer. Sin embargo, de nuevo como p53, hay una compensación. A medida que aumentan los niveles de p16 INK4a en las células madre adultas y las células progenitoras, disminuye su capacidad para reproducirse y así reemplazar el tejido perdido o dañado.
p16 INK4a no es simplemente un reflejo de una célula envejecida, sino que participa activamente en el proceso.
- Los ratones que expresan niveles más altos de lo normal de p16 INK4a muestran una senescencia replicativa más temprana, mientras que
- Los ratones en los que se bloquea la actividad de p16 INK4a continúan reparando el tejido dañado de manera eficiente pero corren un mayor riesgo de contraer cáncer.
- En ratones, eliminar las células senescentes (son altas en p16 INK4a) previene (en ratones jóvenes) y revierte parcialmente (en ratones mayores) algunos de los signos del envejecimiento como las cataratas, y la pérdida de tejido adiposo y masa muscular esquelética.
En humanos, las deleciones y otras mutaciones de p16 INK4a se encuentran en una variedad de cánceres.
Pérdida de heterocigosidad (LOH)
Debido a que los genes supresores de tumores son recesivos, las células que contienen un gen normal y otro mutado —es decir, son heterocigotos— todavía se comportan normalmente. (Excepción: se ha encontrado un gen supresor tumoral ligado al X [WTX]. En los machos, tener solo un cromosoma X, una mutación puntual dañina en WTX o su deleción es todo lo que se necesita para eliminar la supresión tumoral. Las hembras también están en riesgo si la mutación o deleción ocurre en el cromosoma X que no está inactivado).
Sin embargo, existen varios mecanismos que pueden hacer que una célula pierda su gen normal y así esté predispuesta a convertirse en un tumor. Estos pueden resultar en una “pérdida de heterocigosidad” o “LOH”.
Mecanismos de LOH:
- Supresión de
- el alelo normal;
- el brazo cromosómico que contiene el alelo normal;
- el cromosoma completo que contiene el alelo normal (resultando en aneuploidía).
- En mujeres, la inactivación X del cromosoma X que porta el alelo normal.
- Pérdida del cromosoma que contiene el alelo normal seguida de duplicación del cromosoma que contiene el alelo mutado.
- Recombinación mitótica. El estudio de genes supresores tumorales reveló (por primera vez) que el cruce —con recombinación genética— ocurre ocasionalmente en la mitosis (como siempre ocurre en la meiosis).
En #3 y #4, la célula resultante lleva ahora dos copias del gen “malo”. A esto se le llama “reducción a homocigosidad”.
LOH puede funcionar en ambos sentidos.
Cuando la LOH ocurre por recombinación mitótica (proceso #4 anterior), una célula hija se vuelve homocigótica para el alelo mutante pero la otra se vuelve homocigótica para el alelo normal (“tipo salvaje”). Esto no sirve de nada cuando están involucrados genes supresores de tumores pero en otras situaciones puede serlo.
Una rara enfermedad cutánea en humanos llamada ictiosis con confeti es un ejemplo. Es causada por la herencia de una mutación dominante en una de las queratinas (que hacen filamentos intermedios). Al nacer, los lactantes con la enfermedad tienen piel enrojecida uniformemente cuyas células son heterocigóticas para el alelo mutante dominante y un alelo de queratina normal. Pero a medida que pasan los años, aparece un número creciente de parches de piel normal, blanqueada, (como “confeti”). El análisis genómico revela que cada parche se desarrolla a partir de una célula madre cutánea que por recombinación mitótica ha sufrido “reducción a homocigosidad”. En este caso, la célula hija que hereda dos alelos de queratina normales pasa a generar un parche de piel normal. Este trabajo se describe en Choate, K.A., et al. , Ciencia 330:94-97 (1 de octubre de 2010).
La mutación no es la única forma de inactivar los genes supresores de tumores.
Su función también puede ser bloqueada por la metilación de su promotor.
Las células cancerosas a menudo contienen un promotor metilado en un gen supresor de tumores acompañado de
- un promotor bloqueado de manera similar en el otro alelo (produciendo el mismo efecto que #2 anterior);
- una pérdida de ese locus en el otro cromosoma (como la LOH en #1 anterior);
- una mutación inactivante en el otro alelo.
Genes supresores de tumores = antioncogenes

Los genes como RB y p53 también se denominan antioncogenes. Primero se les dio este nombre porque revierten, al menos en cultivo celular, la acción de oncogenes conocidos. Esta imagen (cortesía de Moshe Oren, de Cell 62:671, 1990) muestra placas de Petri las cuales fueron sembradas con el mismo número de células de ratón que habían sido transformadas por dos oncogenes: myc y ras . Muchos de los de la izquierda se han convertido en colonias de células. Sin embargo, las células sembradas a la derecha también contenían el gen supresor tumoral p53. Sólo unos pocos han podido crecer hasta convertirse en colonias.
Virus del Papiloma Humano (VPH)
El nombre anti-oncogén puede ser incluso más apropiado de lo que se pensaba originalmente. Tanto la proteína Rb como la proteína p53 resultan complejarse directamente en la célula con un producto génico de algunos virus del papiloma humano.
Una vez dentro de las células de su huésped, estos virus sintetizan una proteína denominada E7 y otra denominada E6.
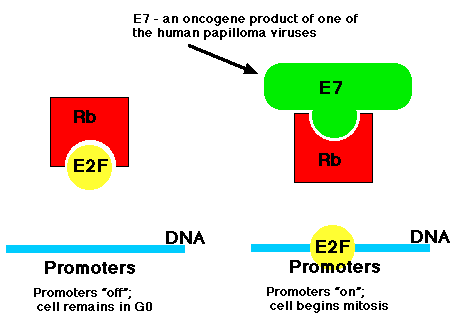
De las >30 cepas de VPH que infectan a humanos, varias, especialmente HVP-16 y HPV-18, han sido implicadas como factor de riesgo de cáncer de cuello uterino y también cánceres de garganta. Su proteína E7 se une a la proteína Rb evitando que se una al factor de transcripción del hospedador E2F.
Resultado: E2F ahora es libre de unirse a los promotores de genes (como c- myc) que hacen que la célula entre en el ciclo celular (derecha). Así, esta versión de E7 es un producto oncogén.
La proteína E6 se une a la proteína p53 dirigiéndola para su destrucción por los proteasomas y eliminando así el bloqueo en la entrada de la célula huésped en el ciclo celular.
Si bien la figura muestra a los promotores “off” como vacíos, ahora está claro que estar “off” implica tanto
- la ausencia de activadores de la transcripción y
- la presencia de represores de transcripción.
Una célula no puede permanecer en G 0 del ciclo celular sin estos represores. Quizás las versiones mutantes de ellos son otra causa de cáncer (las células cancerosas nunca están en G 0).