
4.3: Mecanismos de Catálisis
- Page ID
- 52874

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Una versión imprimible de esta sección está aquí: BiochemFFA_4_3.pdf. Todo el libro de texto está disponible de forma gratuita de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
La magia de las enzimas, como se señaló, está en su capacidad para crear entornos electrónicos propicios para el inicio de una reacción. Hay más mecanismos de reacción de los que podríamos esperar cubrir en un libro como este, y la discusión integral de estos no es nuestro objetivo. En cambio, citaremos algunos ejemplos y entraremos en detalles sobre uno de ellos: el mecanismo de acción de las serina proteasas.
quimotripsina
Comenzaremos con el mecanismo de acción de una enzima - la quimotripsina. Encontrada en nuestro sistema digestivo, la actividad catalítica de la quimotripsina es escindir enlaces peptídicos en proteínas y utiliza la cadena lateral de una serina en su mecanismo de catálisis. Muchas otras enzimas cortantes de proteínas emplean un mecanismo muy similar y se conocen colectivamente como serina proteasas (Figura 4.52).
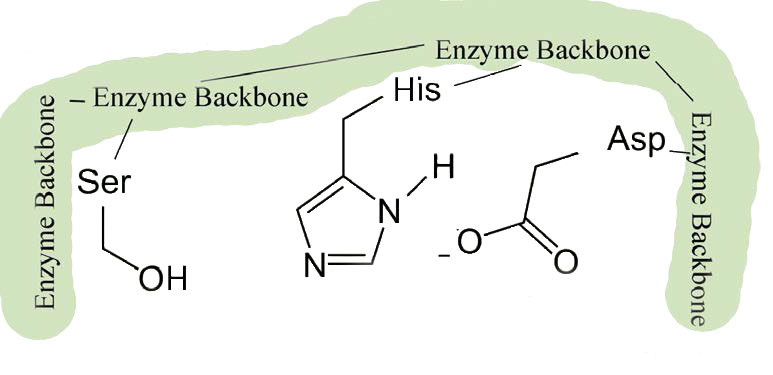
Estas enzimas se encuentran en células procariotas y eucariotas y todas utilizan un conjunto común de tres aminoácidos en el sitio activo llamado tríada catalítica (Figura 4.53). Consiste en ácido aspártico, histidina y serina. La serina se activa en el mecanismo de reacción para formar un nucleófilo en estas enzimas y le da nombre a la clase. Con la excepción del reconocimiento que ocurre en el sitio de unión al sustrato, el mecanismo mostrado aquí para la quimotripsina sería aplicable a cualquiera de las serina proteasas.
Especificidad
Como proteasa, la quimotripsina actúa de manera bastante específica, cortando no todos los enlaces peptídicos, sino solo aquellos que están adyacentes a aminoácidos relativamente no polares en la proteína. Uno de los aminoácidos a los que corta adyacente es la fenilalanina. La acción de la enzima ocurre en dos fases: una fase rápida que ocurre primero y una fase más lenta que sigue. La enzima tiene un sitio de unión al sustrato que incluye una región de la enzima conocida como el bolsillo S1. Pasemos por el mecanismo por el cual la quimotripsina corta adyacente a la fenilalanina.
Encuadernación sustrato
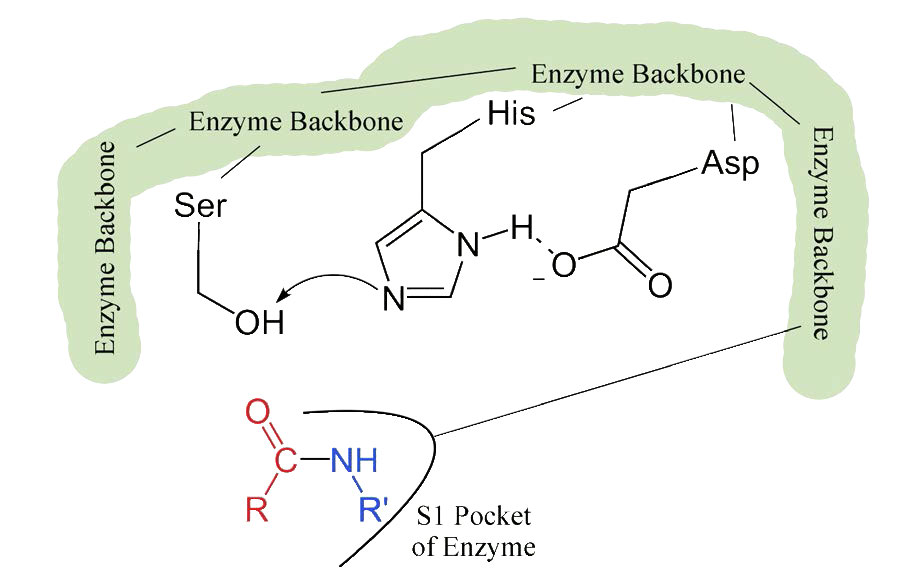
El proceso inicia con la unión del sustrato en el bolsillo S1 (Figura 4.54). El bolsillo S1 en la quimotripsina tiene un orificio hidrofóbico en el que se une el sustrato. Los sustratos preferidos incluirán cadenas laterales de aminoácidos que son voluminosas e hidrófobas, como la fenilalanina. Si una cadena lateral ionizada, como la del ácido glutámico se une en el bolsillo S1, saldrá rápidamente, al igual que el agua evitaría un interior aceitoso.
Cambio de forma en encuadernación
Cuando el sustrato adecuado se une en el bolsillo S1, su presencia induce un cambio cada vez más leve en la forma de la enzima. Este sutil cambio de forma en la unión del sustrato apropiado inicia los pasos de la catálisis. Dado que el proceso catalítico solo comienza cuando se une el sustrato adecuado, esta es la razón por la que la enzima muestra especificidad para cortar en aminoácidos específicos en la proteína diana. Solo los aminoácidos con las cadenas laterales que interactúan bien con el bolsillo S1 inician el giro de las ruedas catalíticas.
Los ligeros cambios de forma implican cambios en el posicionamiento de tres aminoácidos (ácido aspártico, histidina y serina) en el sitio activo conocido como tríada catalítica.
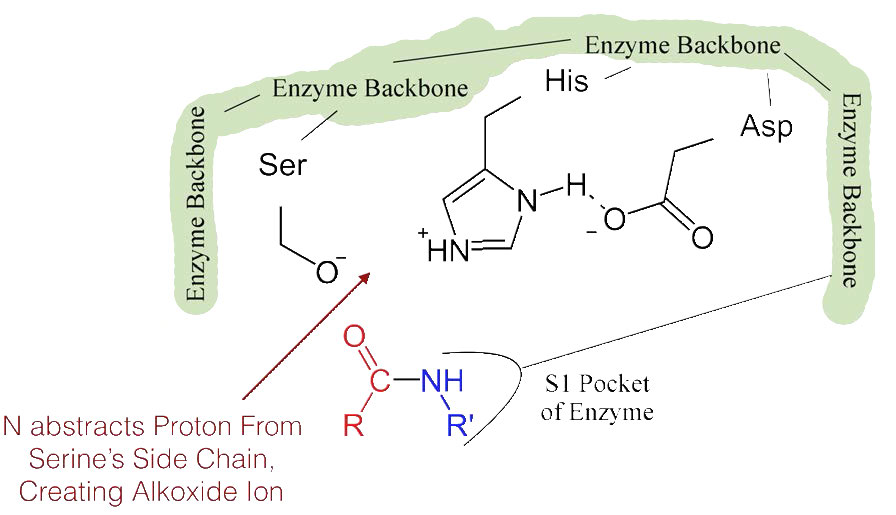
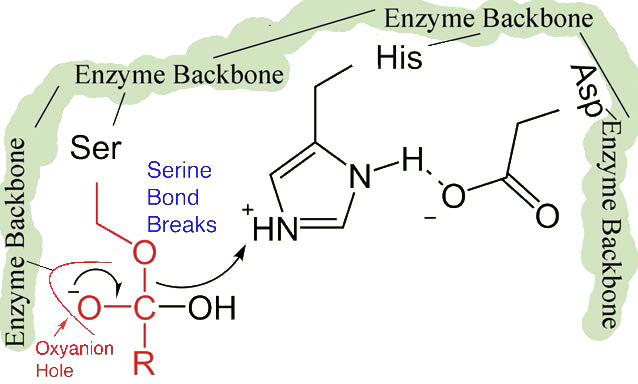
El desplazamiento del ácido aspártico cargado negativamente hacia el anillo de histidina rico en electrones favorece la abstracción de un protón por la histidina del grupo hidroxilo en la cadena lateral de la serina, resultando en la producción de un ion alcóxido muy reactivo en el sitio activo (Figura 4.55).
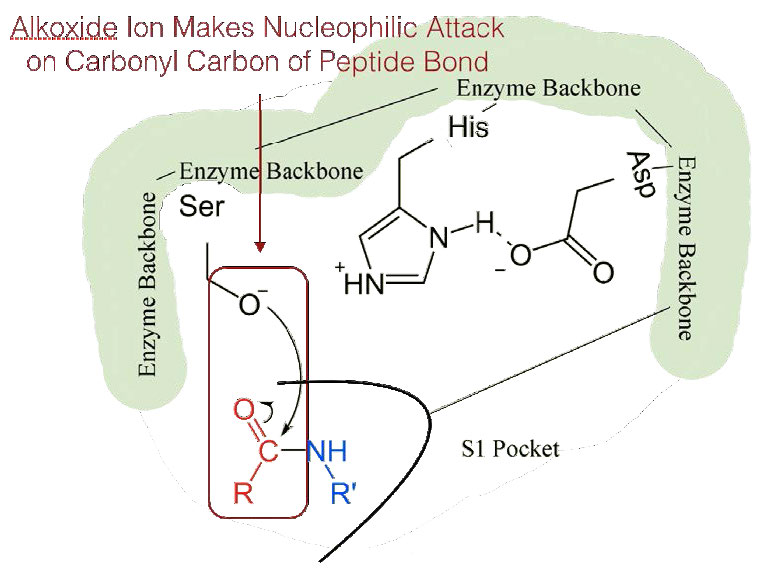
Dado que el sitio activo en este punto también contiene la cadena polipeptídica posicionada con la cadena lateral de fenilalanina incrustada en el bolsillo S1, el ion alcóxido realiza un ataque nucleófilo sobre el enlace peptídico en el lado carboxilo de la fenilalanina que se encuentra en el bolsillo S1 (Figura 4.56). Esta reacción rompe el enlace peptídico (Figura 4.57) y provoca que ocurran dos cosas.
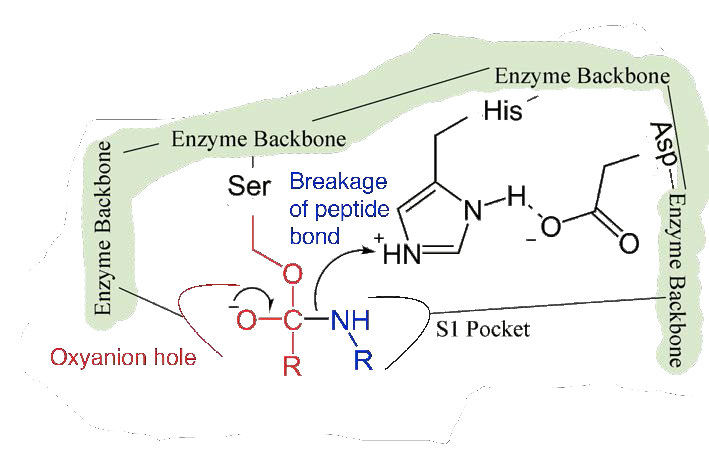
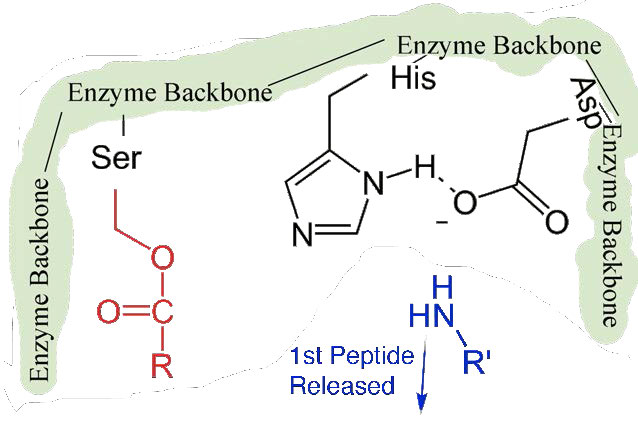
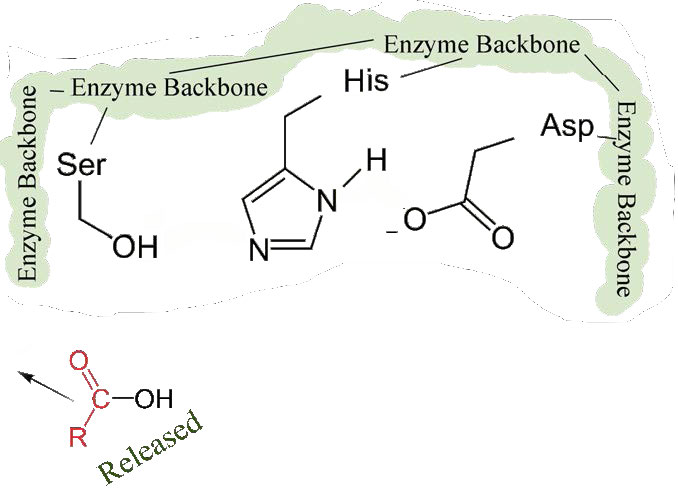
Primero, se libera un extremo del polipéptido original y sale del sitio activo (Figura 4.58). El segundo es que el extremo que contiene la fenilalanina está unido covalentemente al oxígeno de la cadena lateral de la serina. En este punto hemos completado la primera fase (rápida) de la catálisis.
Segunda fase más lenta
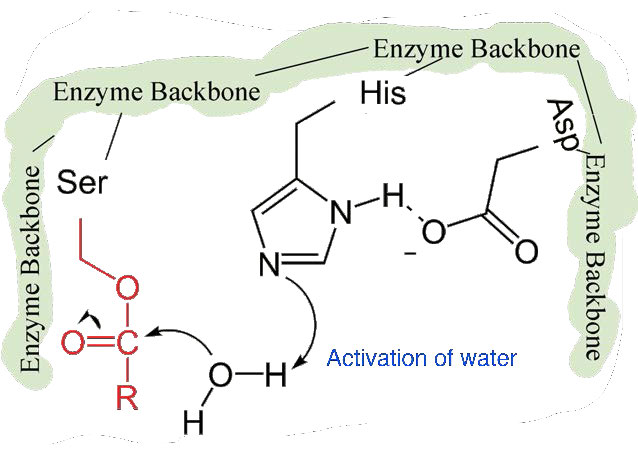
La segunda fase de la catálisis por quimotripsina es más lenta. Requiere que el enlace covalente entre la fenilalanina y el oxígeno de la serina se rompa para que el péptido pueda liberarse y la enzima pueda regresar a su estado original. El proceso inicia con la entrada de agua en el sitio activo. El agua es atacada de manera similar a la de la cadena lateral de serina en la primera fase, creando un grupo hidroxilo reactivo (Figura 4.59) que realiza un ataque nucleófilo sobre el enlace fenilalanina-serina (Figura 4.60), liberándolo y reemplazando el protón sobre la serina. El segundo péptido se libera en el proceso y la reacción se completa con la enzima nuevamente en su estado original (Figura 4.61).
Serina proteasas
La lista de serina proteasas es bastante larga. Se agrupan en dos categorías amplias: 1) las que son similares a la quimotripsina y 2) las que son similares a la subtilisina. Aunque las enzimas de tipo subtilisina y de tipo quimotripsina utilizan el mismo mecanismo de acción, incluida la tríada catalítica, las enzimas no están relacionadas entre sí por secuencia y parecen haber evolucionado de forma independiente. Son, así, un ejemplo de evolución convergente -un proceso donde la evolución de diferentes formas convergen sobre una estructura para proporcionar una función común.
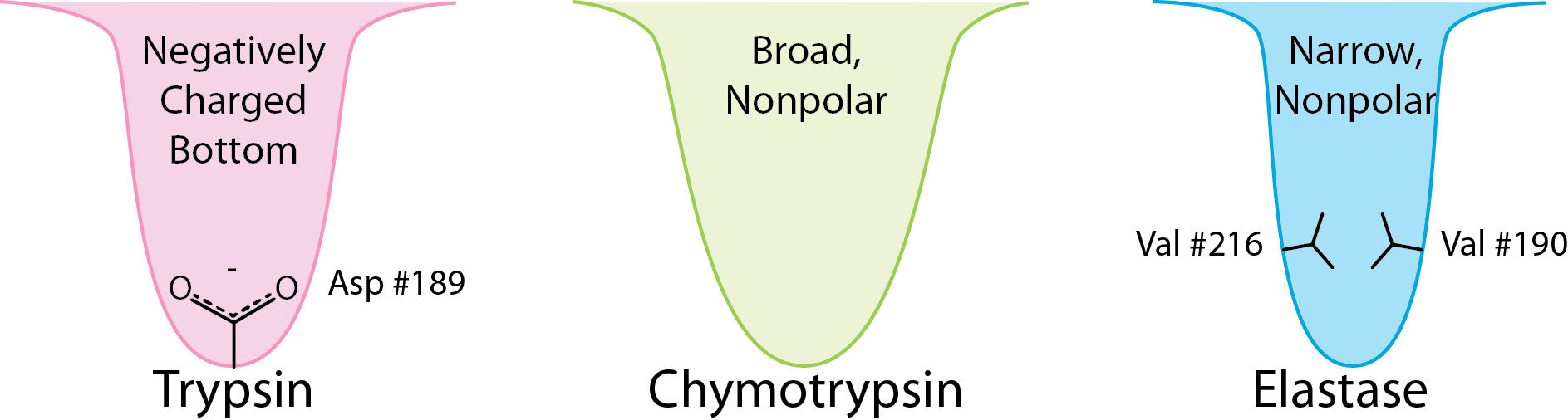
Las enzimas serina proteasa cortan adyacentes a aminoácidos específicos y la especificidad está determinada por el tamaño/forma/carga de la cadena lateral de aminoácidos que encaja en el bolsillo de unión S1 de la enzima (Figura 4.62).
Los ejemplos de serina proteasas incluyen tripsina, quimotripsina, elastasa, subtilisina, peptidasa señal I y nucleoporina. Las serina proteasas participan en muchos procesos fisiológicos, incluyendo la coagulación de la sangre, la digestión, la reproducción y la respuesta inmune.
Proteasas de cisteína
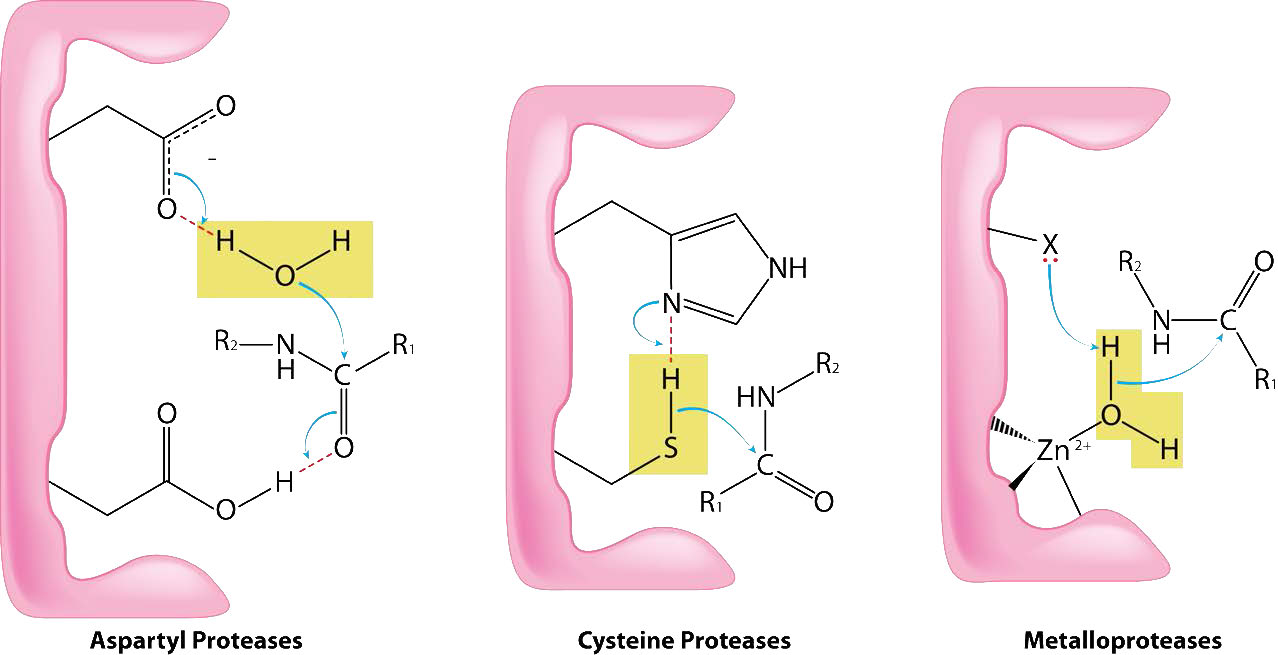
Las cisteína proteasas (también conocidas como tiol proteasas) catalizan la descomposición de las proteínas mediante la escisión de enlaces peptídicos usando un tiol nucleófilo de una cisteína (Figura 4.63). La cisteína se encuentra típicamente en una díada o tríada catalítica que también involucra histidina y (a veces) ácido aspártico (muy parecido a las serina proteasas). El grupo sulfhidrilo de las cisteína proteasas es más ácido que el hidroxilo de las serina proteasas, por lo que no siempre se necesita el ácido aspártico de la tríada.
El mecanismo de acción es muy similar al de las serina proteasas. La unión del sustrato apropiado da como resultado la activación del tiol (eliminación del protón por el grupo histidina). El tiol activado actúa como nucleófilo, atacando el enlace peptídico y provocando que se rompa. Un péptido se libera y el otro péptido se une covalentemente al azufre. La hidrólisis por agua libera el segundo péptido y completa el ciclo. Los ejemplos de cisteína proteasas incluyen papaína, caspasas, proteína hedgehog, calpain y catepsina K.
Caspasas
Las caspasas (cisteína-proteasas aspárticas) son una familia de cisteína proteasas que desempeñan papeles importantes en el organismo. A nivel celular funcionan en apoptosis y necrosis y en el cuerpo, están involucrados en la inflamación y el sistema inmune. La maduración de los linfocitos es uno de esos papeles. Son más conocidos, sin embargo, por su papel en la apoptosis, lo que ha dado lugar a descripciones de las mismas como proteínas “verdugo” o “proteasas suicidas” que desmantelan las células en la muerte celular programada.
Se conocen 12 caspasas humanas. Las enzimas se sintetizan como zimógenos procaspasas con un prodominio y otras dos subunidades. El prodominio contiene regiones que le permiten interactuar con otras moléculas que regulan la actividad de la enzima. Las caspasas vienen en dos formas. Las caspasas iniciadoras, cuando se activan, activan las caspasas efectoras. Las caspasas efectoras escinden otras proteínas en la célula. Las dianas para la acción de escisión de la caspasa efectora incluyen las laminas nucleares (proteínas fibrosas que proporcionan integridad estructural al núcleo), ICAD/DFF45 (un inhibidor de la ADNasa), PARP (áreas señalizadoras donde se necesita reparación del ADN) y PAK2 (regulación apoptótica).
La cascada de activación de caspasa puede activarse por la granzima B (una serina proteasa secretada por células asesinas naturales y células T citotóxicas), receptores de muerte celular y el apoptosoma (estructura proteica grande en células apoptóticas estimuladas por la liberación del citocromo C de las mitocondrias). Cada uno de estos activadores es responsable de activar diferentes grupos de caspasas.
Metaloproteasas
Las metaloproteasas (Figura 4.64) son enzimas cuyo mecanismo catalítico para romper enlaces peptídicos involucra un metal. La mayoría de las metaloproteasas usan zinc como metal, pero algunas usan cobalto, coordinado a la proteína por tres residuos de aminoácidos con un agua lábil en la cuarta posición. Se utilizan una variedad de cadenas laterales: histidina, aspartato, glutamato, arginina y lisina. El agua es la diana de acción del metal que, al unirse al sustrato adecuado, abstrae un protón para crear un grupo hidroxilo nucleófilo que ataca el enlace peptídico, escindiéndolo (Figura 4.64). Dado que el nucleófilo aquí no está unido covalentemente a la enzima, ninguno de los péptidos escindidos termina unido a la enzima durante el proceso catalítico. Los ejemplos de metaloproteasas incluyen carboxipeptidasas, aminopeptidasas, insulinasas y termolisina.
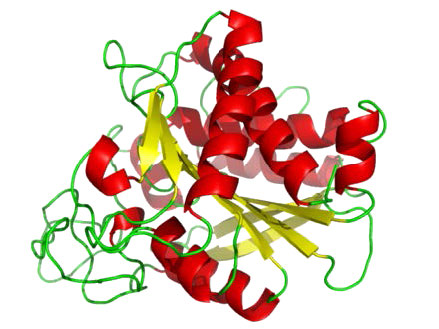
Aspartil proteasas
Como su nombre indica, las aspartil proteasas utilizan ácido aspártico en su mecanismo catalítico (Figuras 4.63 y 4.65). Al igual que las metaloproteasas, las aspartil proteasas activan un agua para crear un nucleófilo para catálisis (Figura 4.65). El agua activada ataca el enlace peptídico del sustrato unido y libera las dos piezas sin necesidad de liberar un intermedio unido, ya que el agua no está unida covalentemente a la enzima. Las aspartil proteasas comunes incluyen pepsina, peptidasa señal II y proteasa del VIH-1.
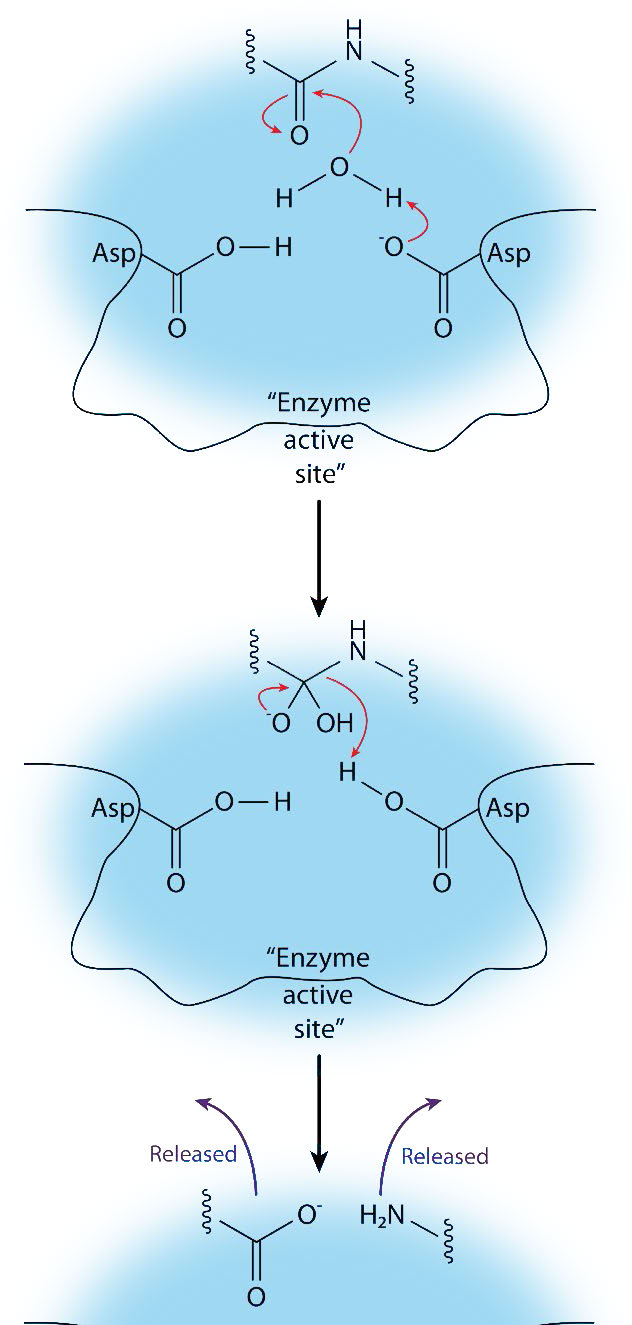
Treonina proteasas
Aunque la treonina tiene un grupo R con un hidroxilo como serina, el mecanismo de acción de esta clase de proteasas difiere algo de las serina proteasas. Hay algunas similitudes. Primero, el hidroxilo de la treonina juega un papel en la catálisis y es actuar como nucleófilo. El nucleófilo se crea, sin embargo, no por una tríada catalítica, sino como resultado del propio grupo α-amina de la treonina que abstrae un protón.
Debido a esto, la treonina nucleofílica en una treonina proteasa debe estar en el extremo n de la enzima. El ataque nucleofílico del enlace peptídico en la proteasa diana da como resultado la rotura del enlace para liberar un péptido y el otro se une covalentemente a la serina, como las serina proteasas. Además, al igual que con las serina proteasas, debe entrar agua para liberar el segundo péptido unido covalentemente para concluir el mecanismo catalítico.
Ejemplos
Los ejemplos de treonina proteasas incluyen las subunidades catalíticas del proteasoma. Algunas acil transferasas (como la ornitina aciltransferasa) han desarrollado el mismo mecanismo catalítico por evolución convergente. Estas últimas enzimas utilizan ornitina en lugar de agua para romper el enlace covalente enzima-sustrato, con el resultado de que el grupo acilo se une a la ornitina, en lugar de agua.
Inhibidores de proteasa
Las moléculas que inhiben la acción catalítica de las proteasas se conocen como inhibidores de proteasas. Estos vienen en una variedad de formas y tienen usos biológicos y medicinales. Muchos inhibidores biológicos son proteínas en sí mismas. Los inhibidores de proteasa pueden actuar de varias maneras, incluyendo como un inhibidor suicida, un inhibidor del estado de transición, un desnaturalizante y como un agente quelante. Algunos trabajan sólo en clases específicas de enzimas. Por ejemplo, la mayoría de las aspartil proteasas conocidas son inhibidas por la pepstatina. Las metaloproteasas son sensibles a cualquier cosa que elimine el metal que requieren para la catálisis. Las metaloproteasas que contienen zinc, por ejemplo, son muy sensibles al EDTA, que quelata el ion zinc.
Una categoría de inhibidores proteicos de proteasas se conoce como las serpinas. Las serpinas inhiben las serinas proteasas que actúan como quimotripsina. 36 de ellas son conocidas en humanos.
Las serpinas son inusuales en actuar uniéndose a una proteasa diana irreversiblemente y experimentando un cambio conformacional para alterar el sitio activo de su diana. Otros inhibidores de proteasa actúan como inhibidores competitivos que bloquean el sitio activo.
Las serpinas pueden ser amplias en su especificidad. Algunos, por ejemplo, pueden bloquear la actividad de las cisteína proteasas. Una de las serpinas biológicas más conocidas es α-1-anti-tripsina (A1AT - Figura 4.66) debido a su papel en los pulmones, donde funciona inhibiendo la proteasa elastasa. La deficiencia de A1AT conduce al enfisema. Esto puede surgir como consecuencia de una deficiencia genética o por fumar cigarrillos. Las especies reactivas de oxígeno producidas por fumar cigarrillos pueden oxidar un residuo crítico de metionina (#358 de la forma procesada) en A1AT, lo que lo hace incapaz de inhibir la elastasa. Desinhibida, la elastasa puede atacar el tejido pulmonar y causar enfisema. La mayoría de las serpinas funcionan extracelularmente. En la sangre, por ejemplo, las serpinas como la antitrombina pueden ayudar a regular el proceso de coagulación.
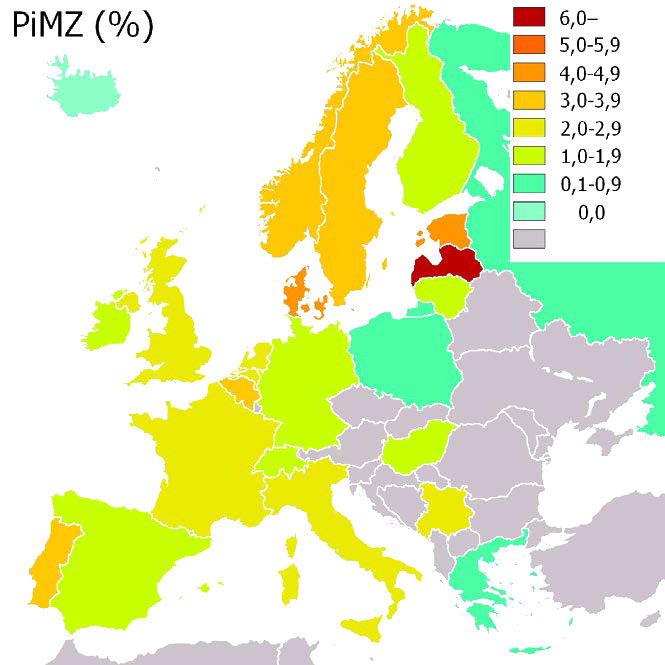
Figura 4.67 - Incidencia de deficiencia de α-1-antitripsina (PiMZ) en Europa en porcentaje. Wikipedia
Antivirales
Los inhibidores de proteasa se utilizan como agentes antivíricos para prohibir la maduración de proteínas virales, comúnmente proteínas de cubierta virales.
Forman parte de los “cócteles” de drogas que se utilizan para inhibir la propagación del VIH en el organismo y también se utilizan para tratar otras infecciones virales, entre ellas la hepatitis C. También se han investigado para su uso en el tratamiento de la malaria y también pueden tener alguna aplicación en terapias anticancerígenas.