
6.3.2: Polipodiopsida
- Page ID
- 58356

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Diferenciar entre helechos, colas de caballo y licófitas.
- Identificar características de brotes vegetativos y reproductivos de Equisetum.
- Identificar características y fases del ciclo de vida del helecho; conocer su ploidía.
- Marque un gametofitos y esporofitos de helecho.
El Polypodiopsida incluye las colas de caballo y helechos. Estas plantas producen hojas con venas ramificadas de tejido vascular llamadas megafilas. Al igual que los licófitos, la reproducción asexual del esporófito se puede lograr a través de rizomas. Las esporas producidas en este grupo se convierten en gametofitos que pueden producir tanto anteridios como archegonia.
Características
- Megafílicas. Las hojas tienen venas ramificadas de tejido vascular. Se cree que las megafilas evolucionaron a partir de tallos ramificados. Cincha rellena los espacios entre las ramas, formando la hoja plana de la hoja. La disposición de las venas foliares (haces vasculares) refleja el patrón de ramificación original de los tallos.
- Rizomas. Propogación asexual del esporofito a través de tallos subterráneos.
- Homosporosas. Las esporas haploides se convierten en gametofitos bisexuales que producen tanto anteridios como archegonia, o son capaces de producir uno u otro, dependiendo de las condiciones.
Equisteum (subclase Equisetidae)
Las colas de caballo son un grupo pequeño con un solo género existente, Equisetum, que tiene alrededor de 30 especies herbáceas diferentes que suelen vivir en hábitats húmedos. El nombre común proviene del patrón característico de ramificación: verticilos o anillos de ramas que surgen de un brote sobre el suelo. Las hojas de estas plantas se han reducido a escamas, y en cambio los tallos segmentados son fotosintéticos. Si miras de cerca los nodos de un brote vegetativo verde, verás que las ramas y las hojas no solo han cambiado de roles, también han cambiado de lugar, con las ramas fotosintéticas emergiendo debajo de las hojas papelosas, no fotosintéticas.
Las colas de caballo suelen crecer en lugares arenosos e incorporan sílice en su epidermis del tallo, lo que le da una superficie abrasiva. Debido a esto, los pioneros estadounidenses usarían esta planta para fregar ollas y sartenes. Así recibió el apodo de “rush de fregado”. El tallo tiene múltiples canales, una característica análoga a los tallos de pastos. Los esporangios están asociados con esporangióforos hexangulares acechados producidos en estrobilos terminales. Dentro de los esporangios, hay eláteres que no son células separadas sino partes de las paredes de las esporas. Los gametofitos suelen ser diminutos y dioicos, pero las plantas mismas son homosporosas: los gametofitos suprimidos más pequeños desarrollan solo anteridios, mientras que los gametofitos más grandes desarrollan solo arquegonia.
Morfología de Gametofitos
Los gametofitos de cola de caballo son reducidos y taloides (figura\(\PageIndex{1}\)). Los gametofitos crecen a partir de homosporas y pueden producir tanto anteridios como archegonia.

Morfología de esporofitos
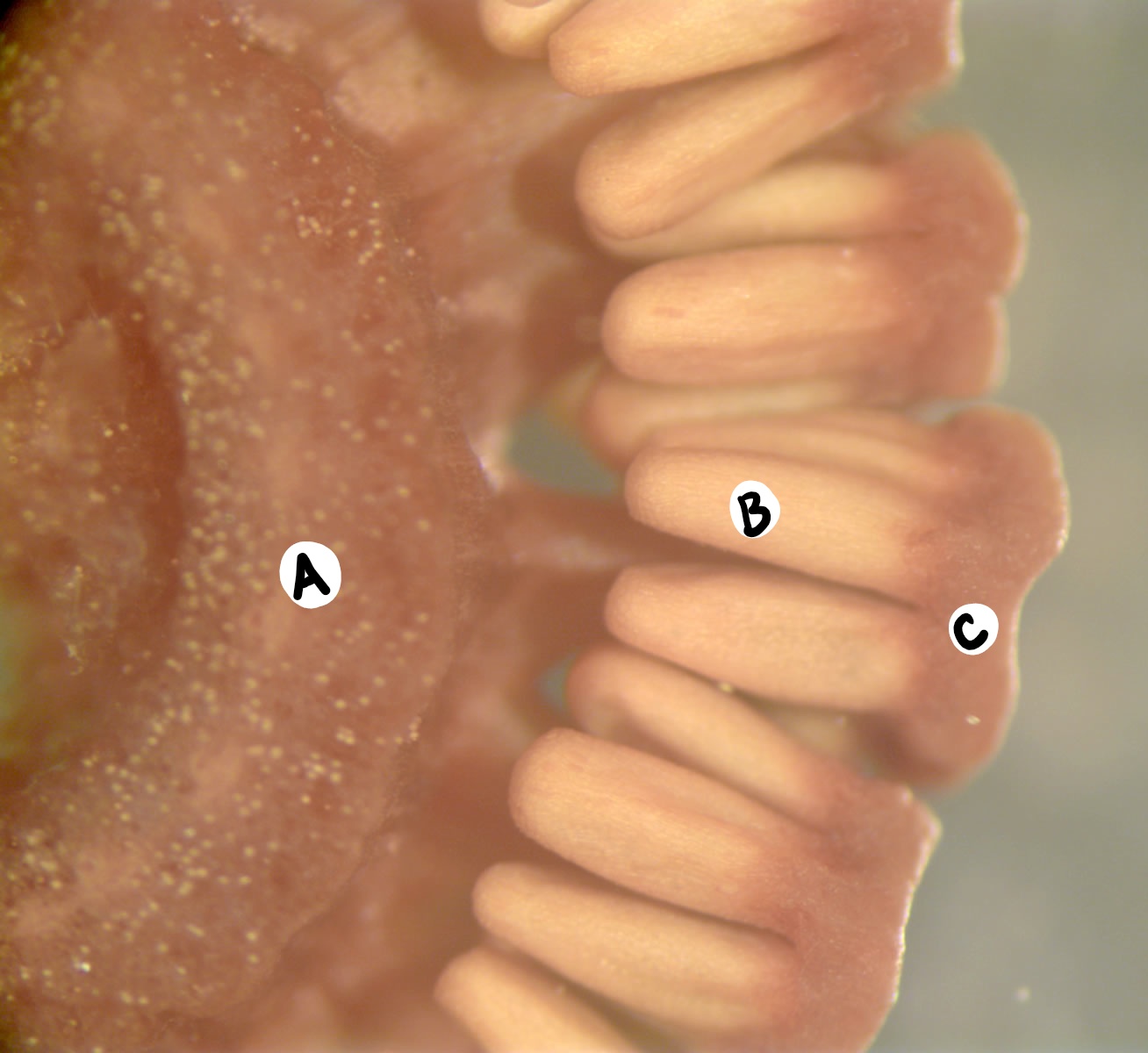
En algunas especies de Equisetum, existen dos tipos diferentes de brotes producidos por el esporófito: los brotes vegetativos que realizan la fotosíntesis y los brotes reproductivos que forman estrobilos y sufren meiosis. En otras especies, el estrobilus se forma en el ápice de un brote fotosintético.
Brotes Vegetativos
En el brote vegetativo, las hojas son oscuras, papelosas y no fotosintéticas. Las ramas son fotosintéticas y se producen en verticilos. Las ramas y hojas emergen en los nodos, separados por regiones del tallo principal llamadas entrenudos. A diferencia de la mayoría de las plantas, las ramas emergen debajo de las hojas en el nodo (Figura\(\PageIndex{2-3}\)). La epidermis de los tallos contiene sílice, la cual tiene una textura abrasiva.


Brotes Reproductivos
Los esporangios se producen en un estrobilo terminal en el brote reproductivo (figura\(\PageIndex{4-5}\)). En algunas especies, este brote reproductivo carece de clorofila y en cambio se alimenta a través del rizoma de brotes vegetativos conectados. Las esporas son fotosintéticas y tienen cuatro brazos higroscópicos llamados elaters.


Video\(\PageIndex{1}\): Este video muestra cómo los eláteres de esporas de Equisetum responden a los cambios en la humedad. Recuperado de YouTube.
Helechos (subclase Polypodiidae)
En la actualidad viven en la tierra unas 15 mil especies de helechos. Muchos de estos se encuentran en los trópicos donde algunos —los “helechos arborescentes ”— pueden crecer a alturas de 40 pies (13 m) o más. Los helechos de las regiones templadas son más pequeños. Por lo general, se encuentran en lugares húmedos y sombreados. Producen rizomas perennes que pueden pasar el invierno. Sus hojas, llamadas frondas debido al crecimiento apical, emergen del rizoma cada primavera como cabezas de violín enrolladas (Figura\(\PageIndex{6}\)). Estas cabezas de violín se abren a través de un proceso llamado vernación circinada, donde los tejidos en crecimiento están protegidos en el centro de la espiral y emergen últimos.
Los verdaderos helechos son megafilos: sus hojas se originaron a partir de ramas aplanadas y tienen venas ramificadas de tejido vascular. Los helechos verdaderos tienen esporangios únicos: leptosporangios (Figura\(\PageIndex{6}\)). Las leptosporangias se originan a partir de una sola célula en una hoja, tienen tallos largos y delgados, y la pared de una capa celular. También se abren activamente: cuando el esporangio madura (se seca), una fila de células con paredes engrosadas en el exterior del esporangio (llamado anillo) se encogerá más lentamente que las células circundantes y finalmente se rompería y liberaría todas las esporas a la vez. Las leptosporangias se agrupan en racimos llamados sori que a menudo están cubiertos con indusia tipo paraguas o bolsillo. Los gametofitos son diminutos y crecen sobre el suelo. Si bien la mayoría de los helechos son homosporosos, algunos géneros de helechos verdaderos (como el helecho acuático Azolla, el tríbol de agua Marsilea y varios otros) son heterosporosos (Figura\(\PageIndex{7}\)).


Los verdaderos helechos son altamente competitivos incluso a las angiospermas. A pesar de su ciclo de vida “primitivo”, tienen múltiples ventajas: habilidades para fotosintetizar en sombra profunda (no están obligados a crecer rápido), para sobrevivir a alta humedad, y para hacer miles de millones de unidades reproductivas (esporas). Los helechos no necesitan gastar sus recursos en flores y frutos, y también son menos vulnerables a los herbívoros vertebrados y plagas de insectos, probablemente porque no los emplean como polinizadores y, por lo tanto, pueden envenenar tejidos contra todos los animales.
Morfología de Gametofitos
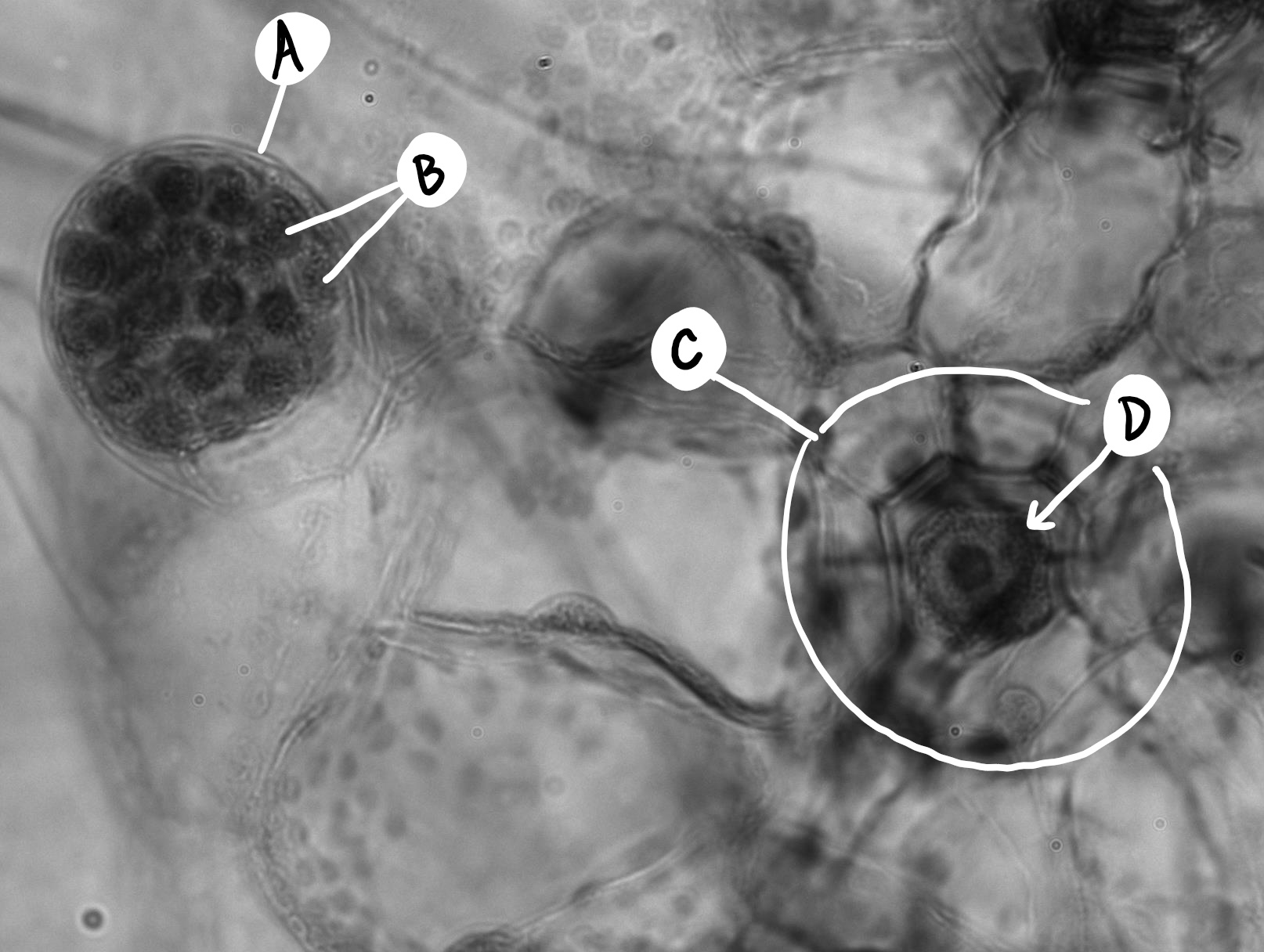
Los gametofitos de helecho son reducidos, taloides y en forma de corazón (Figura\(\PageIndex{8}\)). A menudo se les conoce como protalo o protalio. Los rizoides se producen desde la parte inferior del talo, al igual que en los briófitos. Similar a las colas de caballo, si un gametofito produce anteridios o archegonia puede ser regulado por señales ambientales (Figura\(\PageIndex{9}\)). Cada anteridio produce muchos espermatozoides nadadores, pero la archegonia produce solo un óvulo (Figura\(\PageIndex{10}\)).


Morfología de esporofitos
Los esporofitos de helecho están compuestos por megafilas, a menudo hojas pinnadas compuestas que emergen como cabezas de violinista enrolladas en la primavera. Los esporangios se producen en racimos llamados sori (sorus, singular) en las hojas (Figura\(\PageIndex{11}\)).

La vernación circinada es un término utilizado para describir el desarrollo del fiddlehead de helecho (Figura\(\PageIndex{12}\)) en una fronda. Debido a que las plantas crecen apicamente, es importante proteger los meristemos apicales en los órganos en crecimiento (como hemos visto tanto en yemas axilares como terminales con las escamas protectoras de los cogollos). El violinista es esencialmente una estructura que oculta las puntas crecientes de las hojas. A medida que la fronda se desarrolla, se desdobla gradualmente, soltando las puntas al final.


Un sorus (plural, sori) es un grupo de esporangios, a menudo protegidos por una estructura tipo paraguas llamada indusio a medida que las esporas maduran (Figura\(\PageIndex{13}\)). Algunos sori están protegidos por una extensión de la hoja llamada falsa indusio (Figura\(\PageIndex{14}\)), mientras que otros carecen de cualquier cobertura protectora. Cada esporangio está revestido por una tira inflada de células llamada anillo. Cuando las esporas han madurado, las células en el anillo comienzan a secarse, provocando que las células colapsen y tiren del esporangio abierto, liberando las esporas.
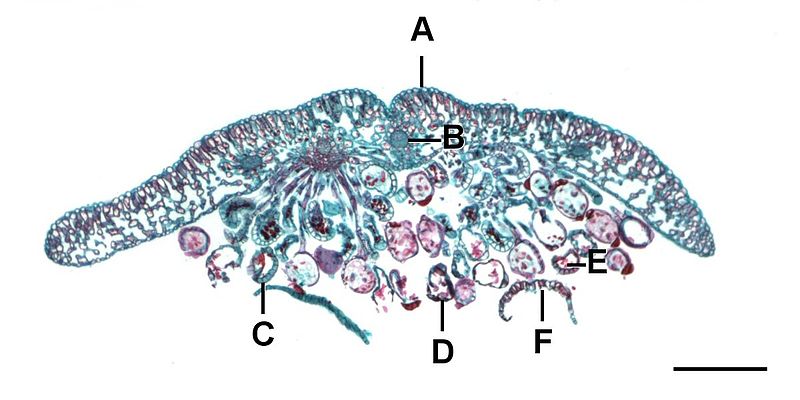

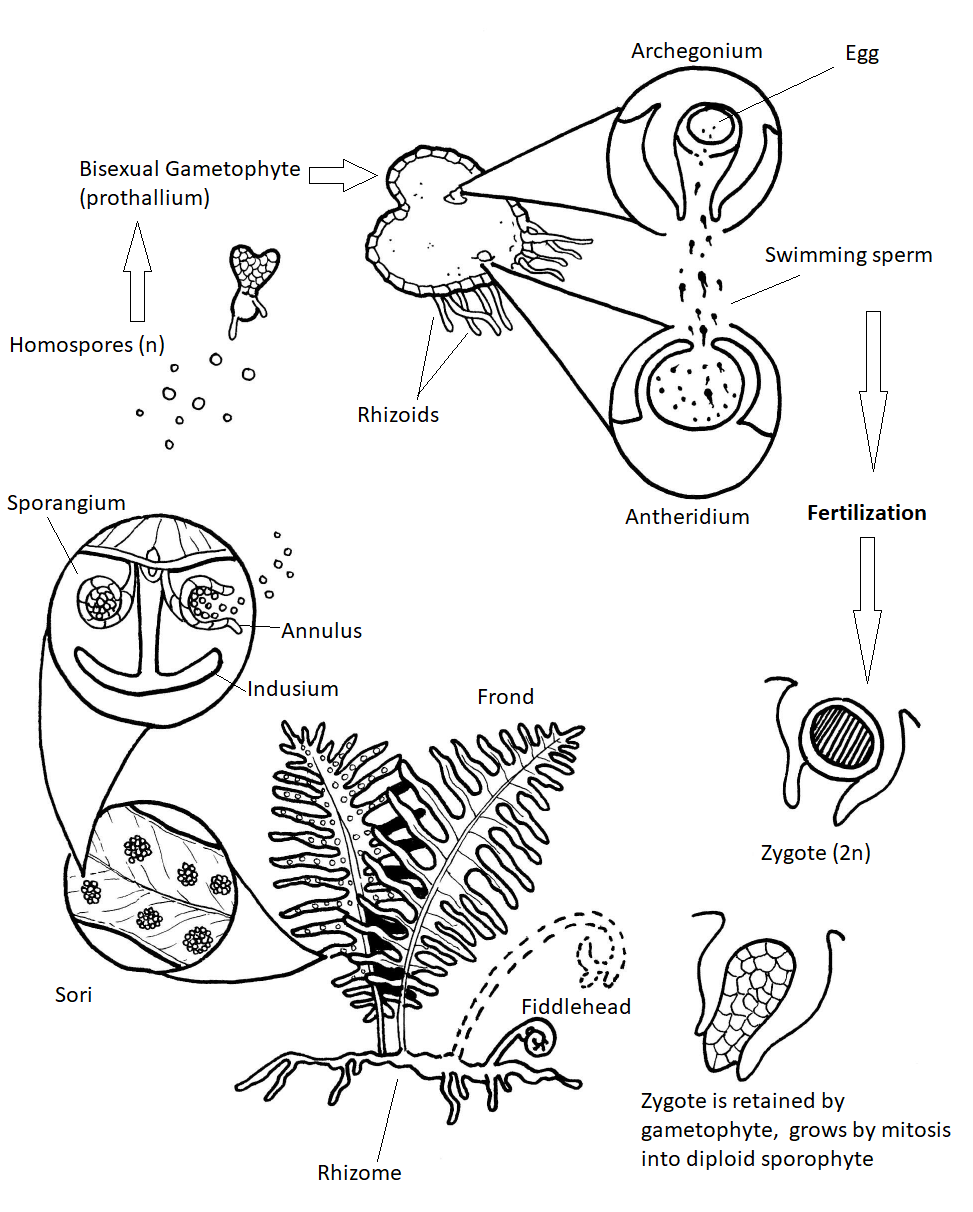
Diagrama de ciclo de vida completo
Los helechos dependen del agua para la dispersión de los espermatozoides, los cuales deben nadar en un archegonio para fertilizar un óvulo (Figura\(\PageIndex{15}\)). Si la humedad es abundante, los espermatozoides nadan hasta la archegonia, generalmente en otro protalo porque los dos tipos de órganos sexuales generalmente no maduran al mismo tiempo en un solo protalo.
Otro método para promover la fertilización cruzada: Las primeras esporas en germinar se desarrollan en protallia con archegonia. Estos protallia secretan una giberelina a su entorno. Esto es absorbido por los protallia más jóvenes y hace que produzcan anteridios exclusivamente.
La fertilización restaura el número diploide e inicia una nueva generación de esporófitos. El esporofito embrionario desarrolla un pie que penetra en el tejido del protalo y permite que el esporofito asegure la nutrición hasta que se vuelva autosuficiente. Aunque es pequeño, el protalo de helecho haploide es una planta autotrófica totalmente independiente. Pronto, el esporofito es nutricionalmente independiente. Es la etapa más grande y de mayor duración del ciclo de vida. Para reproducirse, se forman muchos sori en la parte inferior de las frondas. Dentro de cada esporangio del soro, las células madre de esporas se someten a meiosis produciendo cuatro esporas haploides cada una.
Cuando baja la humedad, las células labiales de pared delgada de cada esporangio se separan, el anillo se endereza lentamente, luego el anillo se rompe hacia adelante expulsando las esporas. Cada una de estas homosporas puede entonces crecer hasta convertirse en un gametofito capaz de producir anteridios y archegonia.
Atribuciones
- Curada y autoría de Maria Morrow utilizando 16.3C Ciclo de Vida de Helecho y 19.1.5 Diversidad y Relaciones Evolutivas de la Biología de Plantas por John. W. Kimball (con licencia CC-BY)
- Evolución de la megafila e información de Salvinia natans agregada por Melissa Ha