
8.2: Síndromes de polinización
- Page ID
- 58448

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Explicar las estrategias de polinización con respecto a la diversidad genética.
- Explicar qué se entiende por el término síndrome de polinización.
- Utilizar características florales para predecir un polinizador (es) de plantas.
En las angiospermas, la polinización se define como la colocación o transferencia de polen de la antera al estigma de la misma flor u otra flor. En las gimnospermas, la polinización implica la transferencia de polen del cono macho al cono hembra. Al transferirse, el polen germina para formar el tubo de polen y el esperma para fertilizar el óvulo.
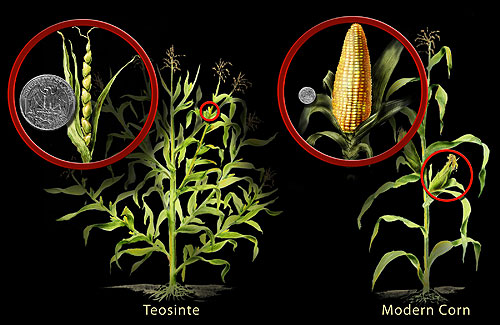
La polinización ha sido bien estudiada desde la época de Gregor Mendel. Mendel realizó exitosamente la autopolinización así como la polinización cruzada en arvejas mientras estudiaba cómo se transmitieron las características de una generación a la siguiente. Los cultivos actuales son el resultado del fitomejoramiento, que emplea la selección artificial para producir los cultivares actuales (término que hace referencia a las variedades cultivadas). Un caso en cuestión es el maíz de hoy, el cual es resultado de años de cría que comenzaron con su antepasado, una hierba llamada teosinte (ver Figura\(\PageIndex{1}\)). El teosinte que originalmente comenzaron a cultivar los antiguos mayas tenía semillas pequeñas, muy diferentes de las mazorcas de maíz y granos regordetes de hoy en día, relativamente gigantes. Curiosamente, aunque estas dos plantas parecen ser completamente diferentes, la diferencia genética entre ellas es minúscula.
.jpg)
.jpg)
Polinización y Diversidad Genética
La polinización toma dos formas: autopolinización y polinización cruzada. La autopolinización ocurre cuando el polen de la antera se deposita sobre el estigma de la misma flor, u otra flor en la misma planta. La autopolinización ocurre en flores donde el estambre y el carpiano maduran al mismo tiempo, y se posicionan de manera que el polen pueda aterrizar sobre el estigma de la flor. Este método de polinización no requiere una inversión de la planta para proporcionar néctar y polen como alimento para los polinizadores, pero da como resultado una menor diversidad genética dentro de la población. La autopolinización suele existir como un “plan B”, en caso de que la polinización cruzada sea, por alguna razón, imposible. En ocasiones, las flores autopolinizadas incluso no se abren; estas flores se llaman cleistogamas.
La polinización cruzada es la transferencia de polen de la antera de una flor al estigma de otra flor en diferentes individuos de la misma especie. Este método se basa en un vector de polen pero da como resultado una mayor diversidad genética.
Las especies vivas se han adaptado para asegurar la supervivencia de su progenie; las que fallan se extinguen. Por lo tanto, se requiere diversidad genética para que en condiciones ambientales cambiantes o de estrés, parte de la progenie pueda sobrevivir. La autopolinización conduce a la producción de plantas con menor diversidad genética, ya que el material genético de la misma planta se utiliza para formar gametos, y eventualmente, el cigoto. Por el contrario, la polinización cruzada —o out-crossing—conduce a una mayor diversidad genética porque el microgametofito y el megagametófito se derivan de diferentes plantas.
Debido a que la polinización cruzada permite una mayor diversidad genética, las plantas han desarrollado muchas formas de evitar la autopolinización. En algunas especies, el polen y el ovario maduran en diferentes momentos. Estas flores hacen que la autopolinización sea casi imposible. Para cuando el polen madura y se ha desprendido, el estigma de esta flor es maduro y sólo puede ser polinizado por el polen de otra flor. Algunas flores han desarrollado características físicas que impiden la autopolinización. La prímola es una de esas flores. Las prímeas han evolucionado dos tipos de flores con diferencias en la longitud de la antera y el estigma: la flor de ojos de pino tiene anteras posicionadas en el punto medio del tubo de polen, y el estigma de la flor de ojos pasantes también se localiza en el punto medio. Los insectos polinizan fácilmente mientras buscan el néctar en el fondo del tubo de polen. Este fenómeno también se conoce como heterostyly. Muchas plantas, como el pepino, tienen flores masculinas y femeninas ubicadas en diferentes partes de la planta, dificultando así la autopolinización. En otras especies, las flores masculinas y femeninas nacen en diferentes plantas (dioicas). Todas estas son barreras para la autopolinización; por lo tanto, las plantas dependen de polinizadores para transferir polen. La mayoría de los polinizadores son agentes bióticos como insectos (como abejas, moscas y mariposas), murciélagos, aves y otros animales. Otras especies de plantas son polinizadas por agentes abióticos, como el viento y el agua.
En resumen, la autoincompatibilidad es un mecanismo que impide la autofertilización en muchas especies de plantas con flores para aumentar la diversidad genética a nivel poblacional. El funcionamiento de este mecanismo de autoincompatibilidad tiene importantes consecuencias para los mejoradores de plantas, ya que inhibe la producción de plantas endogámicas e híbridas.
Genes de incompatibilidad en flores
En las últimas décadas, se han descubierto genes de incompatibilidad, que impiden que el polen germine o crezca en el estigma de una flor, en muchas especies de angiospermas. Si las plantas no tienen genes compatibles, el tubo de polen deja de crecer. La autoincompatibilidad está controlada por el locus S (esterilidad). Los tubos de polen tienen que crecer a través del tejido del estigma y el estilo antes de que puedan ingresar al óvulo. El carpiano es selectivo en el tipo de polen que permite crecer en su interior. La interacción es principalmente entre el polen y las células epidérmicas del estigma. En algunas plantas, como la col, el polen es rechazado en la superficie del estigma, y el polen no deseado no germina. En otras plantas, la germinación del tubo de polen se detiene luego de crecer un tercio de la longitud del estilo, lo que lleva a la muerte del tubo de polen. La muerte del tubo de polen se debe a la apoptosis (muerte celular programada) o a la degradación del ARN del tubo de polen. La degradación es el resultado de la actividad de una ribonucleasa codificada por el locus S. La ribonucleasa es secretada por las células del estilo en la matriz extracelular, que se encuentra junto al tubo de polen en crecimiento.
Síndromes de polinización
Con el tiempo, las angiospermas evolucionaron diferentes morfologías florales, olores y colores que correspondían a su particular vector de polen. Estos conjuntos de características, llamados síndromes de polinización, permiten a los científicos predecir los polinizadores para diferentes plantas. Por ejemplo, las flores en forma de copa suelen ser polinizadas con animales masivos como escarabajos e incluso murciélagos. Las flores en forma de embudo así como las flores labiadas (con labios), están adaptadas a moscas y abejas. Las flores con espolones largos atraen a mariposas y aves (como colibríes o pájaros azucareros). Asimismo, muchos de los vectores del organismo desarrollaron características o comportamientos para especializarse en la polinización de plantas particulares. Esta evolución en tándem hacia el aumento de la especificidad se llama coevolución. Curiosamente, los fenotipos de flores también pueden correlacionarse con otros factores ambientales (por ejemplo, Peach et al. 2020). Para más imágenes de los síndromes de polinización, consulte el Capítulo 14.1 Síndromes de polinización en el Atlas Fotográfico.
Polinización por Decepción
Las orquídeas son flores muy valoradas, con muchas variedades raras (Figura\(\PageIndex{2}\)). Crecen en una variedad de hábitats específicos, principalmente en los trópicos de Asia, América del Sur y Centroamérica. Se han identificado al menos 25 mil especies de orquídeas.

Las flores suelen atraer polinizadores con recompensas alimentarias, en forma de néctar. Sin embargo, algunas especies de orquídeas son una excepción a este estándar: han evolucionado diferentes formas de atraer a los polinizadores deseados. Utilizan un método conocido como engaño alimentario, en el que se ofrecen colores brillantes y perfumes, pero no comida. Anacamptis morio, comúnmente conocida como la orquídea de alas verdes, tiene flores de color púrpura brillante y emite un fuerte aroma. El abejorro, su polinizador principal, se siente atraído por la flor debido al fuerte olor —que suele indicar alimento para una abeja— y en el proceso, recoge el polen para ser transportado a otra flor.
Otras orquídeas utilizan el engaño sexual. Chiloglottis trapeziformis emite un compuesto que huele igual que la feromona emitida por una avispa hembra para atraer avispas macho. La avispa macho se siente atraída por el aroma, aterriza sobre la flor de la orquídea y, en el proceso, transfiere polen. Algunas orquídeas, como la orquídea martillo australiana, utilizan el aroma y el engaño visual en otra estrategia de engaño sexual para atraer avispas. La flor de esta orquídea imita la apariencia de una avispa hembra y emite una feromona. La avispa macho intenta aparearse con lo que parece ser una avispa hembra, y en el proceso, recoge polen, que luego transfiere al siguiente mate falsificado.
Abejas
Las abejas son quizás el polinizador más importante de muchas plantas de jardín y árboles frutales más comerciales (Figura\(\PageIndex{3}\)). La mayoría de las abejas que la gente nota son las abejas sociales, abejorros y abejas melíferas, aunque hay miles de especies de abejas solitarias que tienen papeles esenciales de polinización. Las abejas tienen pelos ramificados que cubren sus cuerpos, lo que las convierte en excelentes polinizadores, ya que es probable que más granos de polen sean capturados en sus superficies externas y transferidos a otras plantas.
Las abejas recolectan polen y néctar ricos en energía para su supervivencia y necesidades energéticas, por lo que las flores que han coevolucionado con las abejas contienen tanto néctar como polen. Dado que las abejas no pueden ver el color rojo, las flores polinizadas por abejas suelen tener tonos de azul, amarillo, violeta u otros colores. Visitan flores que están abiertas durante el día, son de colores brillantes, tienen un fuerte aroma o aroma, con una zona de aterrizaje de algún tipo, típicamente con la presencia de un guía de néctar. Una guía de néctar puede incluir regiones en la flor con pigmentos que son visibles solo para las abejas, y no para los humanos (como UV); ayuda a guiar a las abejas al centro de la flor, haciendo así que el proceso de polinización sea más eficiente. El polen se adhiere al pelo borroso de las abejas, y cuando la abeja visita otra flor, parte del polen se transfiere a la segunda flor.


Recientemente, las poblaciones de abejas melíferas (una especie europea introducida) han estado en declive debido a una variedad de factores, entre ellos los pesticidas (como los neonicotinoides) y los ácaros parásitos. Debido a que las abejas solitarias no son operadas por la industria comercial, los impactos en estas poblaciones han sido menos documentados. Sin embargo, la disminución de las poblaciones de abejas tendrá impactos extremadamente importantes en nuestra seguridad alimentaria, ya que las abejas son utilizadas para polinizar muchos cultivos alimentarios.
Las avispas también son importantes polinizadores de insectos, y polinizan muchas especies de higos.
Moscas
Muchas especies de moscas son polinizadores importantes. Estas moscas suelen verse y comportarse de manera muy parecida a las abejas, desempeñando papeles importantes en la polinización. Sin embargo, dado que estas flores generalmente se clasificarían bajo un síndrome de polinización de abejas, en su lugar discutiremos una segunda categoría de mosca para el síndrome de polinización de la mosca. Estos insectos (moscas y a veces escarabajos carroñeros) normalmente no buscan una flor, sino un cadáver o algún estiércol para alimentarse o poner huevos en ellos. Algunas flores han evolucionado para engañar a las moscas hacia la polinización emitiendo un olor a descomposición o un olor a carne podrida. Estas flores, que producen néctar, suelen tener colores apagados, como el marrón o el morado. A menudo presentan algún tipo de característica similar a una trampa, por lo que cuando una mosca visita, tiene que rebotar un poco dentro de la flor antes de irse (esto aumenta la posibilidad de deposición de polen, tanto sobre la flor como sobre la marcha). Los ejemplos incluyen las flores del cadáver (Amorphophallus y Rafflesia) y dragon arum (Dracunculus).

Mariposas y polillas
Las mariposas, como la monarca, polinizan muchas flores de jardín y flores silvestres, que suelen aparecer en racimos. Estas flores son de colores brillantes, tienen una fuerte fragancia, están abiertas durante el día y tienen guías de néctar para facilitar el acceso al néctar. El polen es recogido y transportado en las extremidades de la mariposa.
Las polillas, por otro lado, polinizan las flores al final de la tarde y la noche. Las flores polinizadas por las polillas son pálidas o blancas y son planas, permitiendo que las polillas aterricen. Un ejemplo bien estudiado de una planta polinizada por polillas es la planta de yuca, que es polinizada por la polilla de la yuca. La forma de la flor y la polilla se han adaptado de tal manera que permiten una polinización exitosa. La polilla deposita polen sobre el estigma pegajoso para que la fertilización ocurra más tarde. La polilla hembra también deposita óvulos en el ovario. A medida que los huevos se convierten en larvas, obtienen alimento de la flor y semillas en desarrollo. Así, tanto el insecto como la flor se benefician mutuamente en esta relación simbiótica. La polilla del gusano del maíz y la planta de Gaurá tienen una relación similar (Figura\(\PageIndex{5}\)).

murciélagos
En los trópicos y desiertos, los murciélagos suelen ser polinizadores de flores nocturnas como el agave, la guayaba y la gloria matutina. Las flores suelen ser grandes y blancas o de color pálido; así, se pueden distinguir del entorno oscuro por la noche. Las flores tienen una fragancia fuerte, afrutada o almizclada y producen grandes cantidades de néctar. Son naturalmente grandes y de boca ancha para acomodar la cabeza del murciélago (Figura\(\PageIndex{6}\)). A medida que los murciélagos buscan el néctar, sus rostros y cabezas se cubren de polen, que luego se transfiere a la siguiente flor.

Pájaros
Muchas especies de aves pequeñas, como el colibrí (Figura\(\PageIndex{7}\)) y las aves sol, son polinizadores para plantas como orquídeas y otras flores silvestres. Las flores visitadas por las aves suelen ser robustas y están orientadas de tal manera que permiten que las aves permanezcan cerca de la flor sin que sus alas se enreden en las flores cercanas, como colgando (colgante). La flor típicamente tiene una forma curvada y tubular, lo que permite el acceso para el pico del ave. Las flores de colores brillantes e inodoras que están abiertas durante el día son polinizadas por las aves. Como un ave busca néctar rico en energía, el polen se deposita en la cabeza y el cuello del ave y luego se transfiere a la siguiente flor que visita. Se sabe que los botánicos determinan el rango de plantas extintas recolectando e identificando polen de especímenes de aves de 200 años de edad del mismo sitio.

Viento
La mayoría de las especies de coníferas, y muchas angiospermas, como pastos, arces y encinos, son polinizadas por el viento. Los conos de pino son marrones y sin perfume, mientras que las flores de las especies de angiospermas polinizadas por el viento suelen ser verdes, pequeñas, pueden tener pétalos pequeños o nulos, y producen grandes cantidades de polen. A diferencia de las típicas flores polinizadas por insectos, las flores adaptadas a la polinización por el viento no producen néctar ni aroma. En las especies de angiospermas polinizadas por el viento, las anteras suelen ser grandes y cuelgan de la flor, y, a medida que sopla el viento, el polen ligero se transporta con ella (Figura\(\PageIndex{8}\)). Las flores suelen emerger temprano en la primavera, antes que las hojas, para que las hojas no bloqueen el movimiento del viento. El polen se deposita sobre el estigma plumoso expuesto de la flor (Figura\(\PageIndex{9}\)).


Agua
Algunas plantas, como el pasto marino australiano y las malas hierbas de los estanques, son polinizadas por el agua. El polen flota sobre el agua, y cuando entra en contacto con la flor, se deposita dentro de la flor.
Cuadro Resumen de Síndromes de Polinización
El cuadro\(\PageIndex{1}\) resume los síndromes de polinización descritos en las categorías anteriores.
| Síndrome de polinización | Color | Estructura | Aroma | Néctar o Polen |
|---|---|---|---|---|
| Viento | Sin brillo, perianto a menudo ausente o reducido | Grandes estigmas plumosos, anteras grandes | Ninguno | Sin néctar, grandes cantidades de polen |
| Pájaros | Rojos y rosas | A menudo tubular o ahuecado | Ninguno | Mucho néctar oculto, polen moderado |
| Abejas | Morados, azules, amarillos, blancos, UV | Plano y poco profundo o tubular, con área de aterrizaje | Dulce, fresco, suave | Polen a menudo pegajoso y perfumado, néctar generalmente presente |
| murciélagos | Blanco, verde opaco o morado | A menudo en forma de cuenco o colgante, anteras sobresaliendo | Mohoso o afrutado, fuerte, emitido por la noche | Mucho néctar oculto |
| Polillas | Blanco, rosa pálido o morado | A menudo tubular o ahuecado, sin plataforma de aterrizaje | Fuerte y dulce, emitido por la noche | Mucho néctar oculto, polen limitado |
| Mariposas | Colores brillantes | Tubular, con plataforma de aterrizaje ancha | Desmayo, fresco | Mucho néctar oculto, polen limitado |
| Moscas | Rojo oscuro, morado, marrón | Poco profundo, embudo o tipo trampa | Pútrido, putrefacto | Sin néctar, polen moderado |
Atribuciones
Comisariada y autoría de Maria Morrow utilizando las siguientes fuentes:
- 32.2 Polinización y Fertilización de Biología 2e por OpenStax (licenciado CC-BY). Accede gratis en openstax.org.
- 8.2 La flor y el fruto de la introducción a la botánica de Alexey Shipunov (dominio público)