
11.3: Vástago Secundario
- Page ID
- 58725

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Comparar el origen y la función del cambium vascular y el cambium del corcho.
- Define la corteza y distinga entre corteza interna y externa.
- Explicar la producción de madera y relacionarla con los anillos anuales.
- Distinguir entre duramen y albura.
- Distinguir entre madera coníferas y madera dura
- Identificar las características externas de las ramitas de invierno.
El crecimiento primario ocurre cuando un tallo aumenta de longitud como resultado de la división celular en el meristema apical del brote. El crecimiento secundario se caracteriza por un incremento en el grosor o circunferencia de la planta, y es causado por meristemos secundarios de división celular. Las plantas herbáceas en su mayoría experimentan crecimiento primario, con apenas crecimiento secundario o aumento de espesor. El crecimiento secundario o “madera” se nota en las plantas leñosas; ocurre en algunas eudicoteñas, pero ocurre muy raramente en monocotiledóneas.
Meristemos Secundarios (Meristemos Laterales)
Dos meristemos secundarios (meristemos laterales) son responsables del crecimiento secundario: el cambium vascular y el cambium de corcho (Figura\(\PageIndex{1}\)).
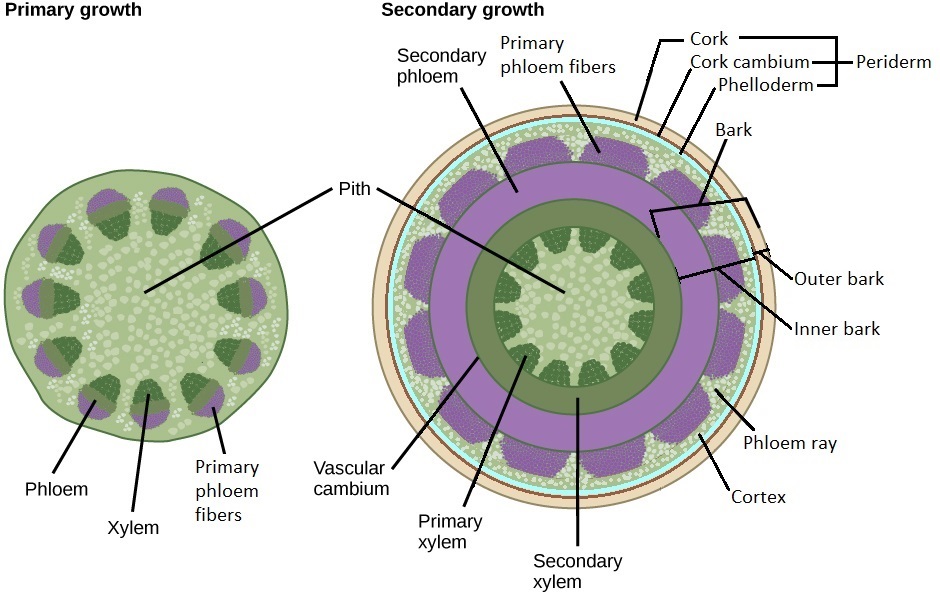
El cambium vascular produce tejido vascular secundario. Las iniciales fusiformes son las células del cámbium vascular que se dividen para producir xilema secundario internamente y floema secundario externamente. Las iniciales de los rayos son las células del cámbium vascular que producen rayos vasculares (rayos de xilema y rayos de floema). Se trata de bandas de parénquima que son perpendiculares a las capas concéntricas de xilema y floema (Figura\(\PageIndex{2}\)). Funcionan en almacenamiento, produciendo compuestos secundarios (moléculas utilizadas por la planta que no son partes esenciales del metabolismo), y transportando materiales entre el xilema y el floema. A medida que el tallo secundario se espesa, los rayos del floema se espesan externamente (convirtiéndose en forma de cuña) para acomodar el diámetro creciente. Si bien el cámbium vascular es técnicamente solo una capa celular de una sola capa, se parece a las capas de células que lo rodean (que recientemente se dividió para producir), y esta región entera a veces se llama el cambium vascular como resultado.
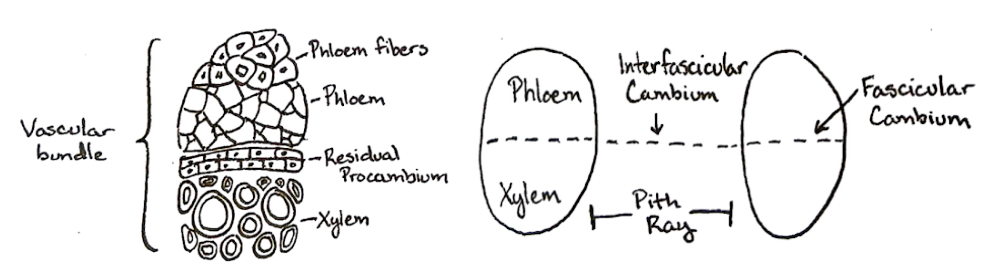
El cambium vascular surge de células madre dentro y entre los haces vasculares en algunos silenosteles y eusteles. Dentro de los haces vasculares, tales células madre (específicamente, células procambiales) forman el cambium fascicular. En las regiones interfasciculares entre haces vasculares se encuentra el cambium interfascicular (Figura\(\PageIndex{3}\)). El cámbium fascicular y el cambium interfascicular finalmente forman el cámbium vascular. En contraste, el cambium vascular en las raíces surge del procámbium y periciclo.
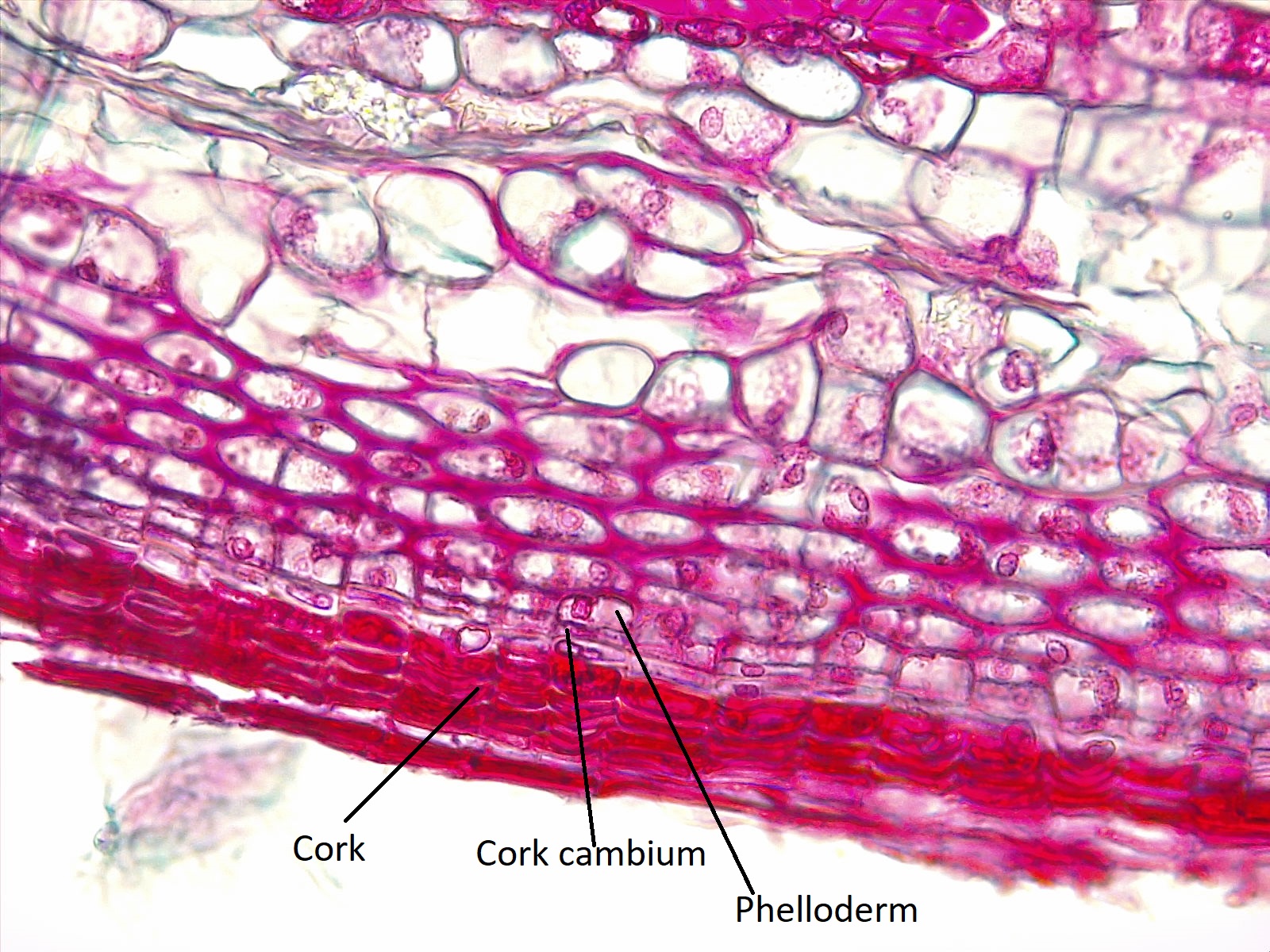
El cambium de corcho se divide para producir felodermo internamente y corcho externamente. Juntos, el felodermo, el cámbium del corcho y el corcho forman el peridermo, el tejido dérmico del cuerpo secundario de la planta (figura\(\PageIndex{4}\)). El primer cambium de corcho producido por un tallo surge de la corteza, pero los posteriores cambios de corcho son producidos por las células del parénquima del floema secundario. (En contraste, el cambium del corcho surge del periciclo en las raíces.)
Las palmeras, que son monocotiledóneas, no tienen meristemos secundarios y madera verdadera. Algún engrosamiento ocurre en una palma pero esto ocurre en la base del árbol, como resultado del crecimiento de raíces adventicias. Las palmas también pueden tener un crecimiento secundario difuso que es la división y agrandamiento de algunas células del parénquima. Estos procesos no compensan el crecimiento general de la planta, y las palmas frecuentemente son más gruesas en la parte superior que en la parte inferior. Otra monocotiledónea, el árbol de sangre de dragón (Dracaena), tiene un crecimiento secundario anómalo, que emplea cambium pero este cambium no forma el anillo estable.
Peridermo y Corteza
Al final del primer año de crecimiento del tallo secundario, el peridermo reemplaza a la epidermis, pero se conservan la corteza y la médula. En contraste, las raíces que experimentan crecimiento secundario no tienen pitos para empezar, y la corteza se pierde durante el crecimiento secundario. Al igual que la epidermis, la mayor parte del peridermo no es permeable al vapor de agua, dióxido de carbono y oxígeno gaseoso. Esto se debe a la suberina cerosa que llena las células de corcho, las cuales están muertas en la madurez. Sin embargo, el intercambio de gases con el ambiente es posible en lenticelas, regiones elevadas del peridermo con muchos espacios de aire intercelulares (Figura\(\PageIndex{5-6}\)). Para producir lenticelas, algunas células de corcho cambium se dividen y crecen mucho más rápido, lo que finalmente romperá el peridermo abierto. Los tallos leñosos no realizan intercambio regular de gases como lo hacen los tallos primarios al abrir y cerrar los estomas, pero las plantas leñosas aún tienen hojas con altas densidades de estomas para regular el intercambio de gases.


La corteza consiste en todas las capas de tejido externas al cámbium vascular. Protege a la planta contra daños físicos y ayuda a reducir la pérdida de agua. En un tallo de un año de interior a exterior, este sería el floema secundario, las fibras primarias del floema, la corteza, el felodermo, el corchete cambium y el corcho. El cámbium de corcho divide la corteza interna y externa. La corteza interna lo es todo dentro del cambium de corcho. La corteza exterior es el cambium del corcho y todo lo externo a él (Figura\(\PageIndex{7}\)).
Un tallo secundario en última instancia produce múltiples capas de peridermo. La corteza interna en un tallo más viejo consiste así en el floema secundario más nuevo y el felodermo más nuevo. Solo el floema conductor de la corteza interna contiene células vivas y transporta materiales mientras que el floema no conductor de la corteza interna contiene células muertas que se utilizan para el almacenamiento. A medida que el tallo secundario envejece, las capas viejas del floema secundario se empujan externamente y se trituran, con la excepción de las fibras del floema, que tienen paredes celulares engrosadas. La corteza externa en un tallo más viejo sería el más nuevo cambium de corcho, el corcho más nuevo y las capas concéntricas de floema viejo y peridermo viejo. Si los peridermos múltiples forman círculos perfectos, la corteza es lisa. Más a menudo, el peridermo múltiple no se superponen de manera uniforme, lo que resulta en corteza áspera con escamas.

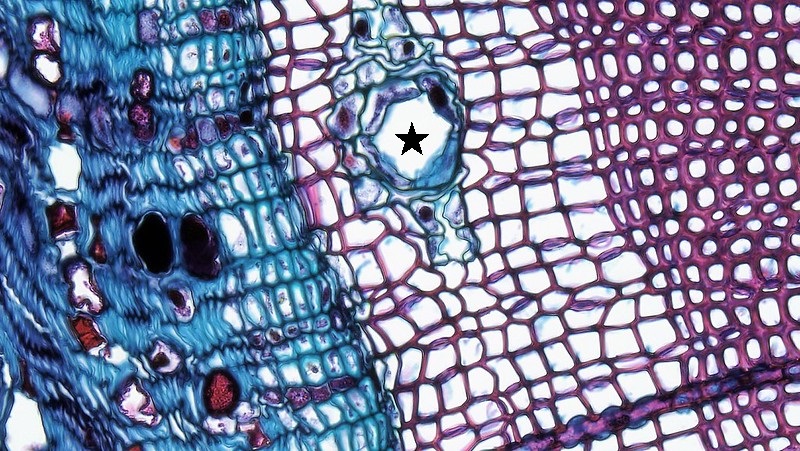
Madera (xilema secundario)
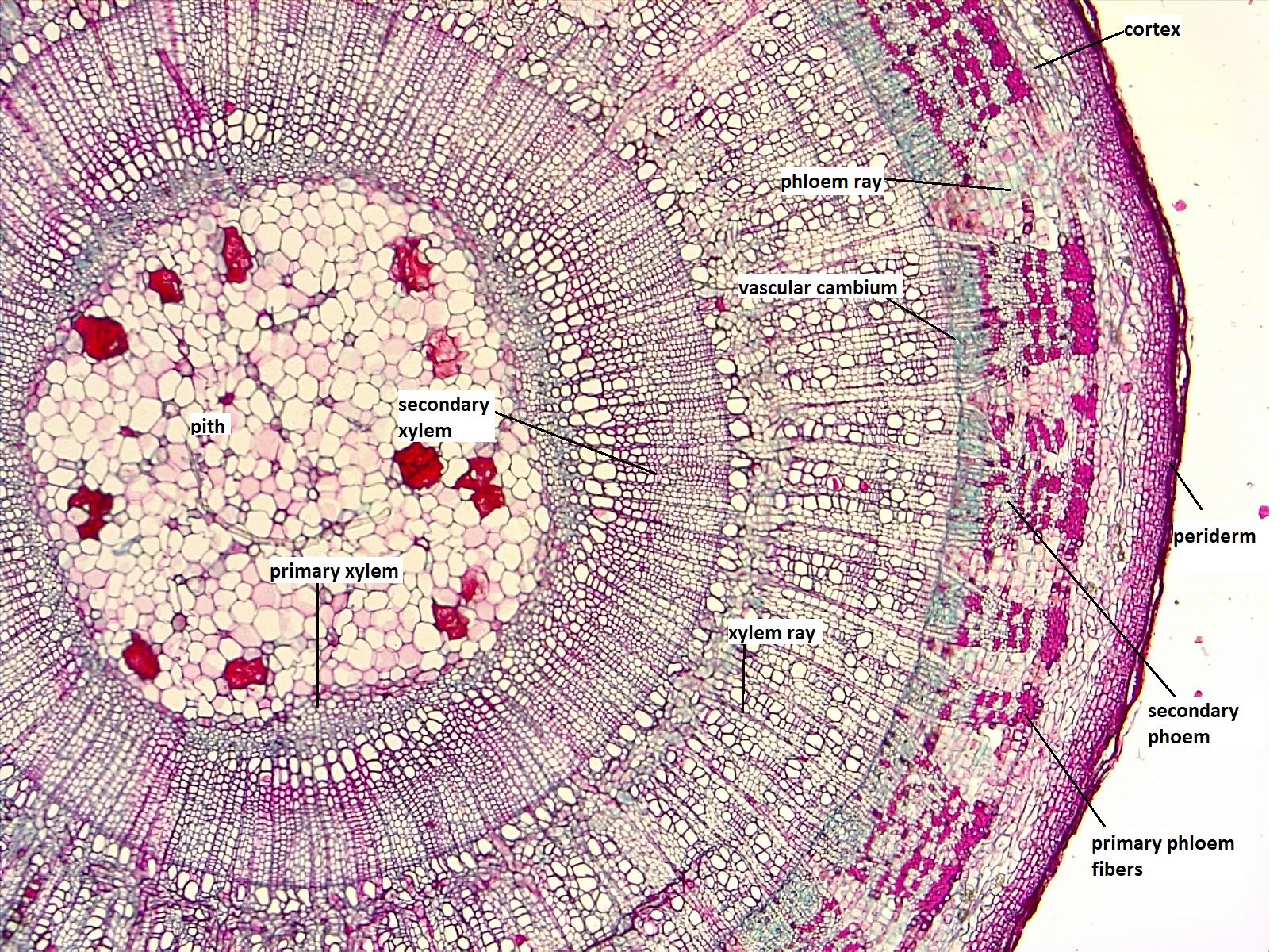
La madera consiste en el xilema secundario producido por el cámbium vascular (Figura\(\PageIndex{8}\)). A diferencia del floema, las viejas capas de xilema secundario se retienen y no se trituran fácilmente. Sin embargo, el xilema secundario más antiguo (cerca del centro del tallo secundario) ya no conduce agua. Se trata del duramen, que almacena diversos compuestos y aparece más oscuro que la madera circundante. Para bloquear el flujo de agua en el duramen, las plantas utilizan tílides elemento buque “tapones”, que también ayudan a controlar el funcionamiento invernal de los vasos. Una tilosa se forma cuando una pared celular del parénquima crece en el elemento traqueario; parecen burbujas. La albura rodea el duramen, es de color más claro, y consiste en el xilema conductor, que se produjo más recientemente (Figura\(\PageIndex{7}\)).

En la primavera de las regiones templadas, el cámbium vascular produce elementos traqueares anchos (las células conductoras del xilema, ya sean elementos vasculares o traqueides). Estos transportan grandes volúmenes de agua, la cual es abundante debido a las lluvias primaverales. Durante el verano, el cambium vascular produce elementos traqueares estrechos como resultado de la menor disponibilidad de agua. En el invierno, la actividad del cambium vascular es baja. Se reanuda la primavera siguiente produciendo nuevamente los amplios elementos traquearios de la madera temprana (madera de primavera), que contrastan claramente con la madera tardía adyacente (madera de verano) del año anterior. La madera temprana parece más ligera y es menos densa que la madera tardía. Cada año de producción de madera es así visible en una sección transversal de un tallo leñoso porque consiste en una capa clara y una capa oscura. Estos se denominan anillos anuales (anillos de árboles; Figura\(\PageIndex{9-10}\)) y pueden ser utilizados para determinar la edad de un árbol o rama a través del estudio de la dendrocronología. Además, los anillos anuales gruesos indican años húmedos y los anillos anuales delgados indican años secos.


Algunos árboles (como encinos, Quercus) tienen grandes elementos de vaso que se encuentran principalmente en la madera temprana; este patrón se conoce como anillo poroso (Figura\(\PageIndex{10}\)). Los grandes elementos de vasos de otros árboles (como olmo, Ulmus) ocurren de manera más uniforme tanto en la madera temprana como tardía. Este patrón se conoce como madera porosa difusa: con grandes elementos de vaso tanto en maderas tempranas como tardías. (Las especies porosas difusas aún producen anillos anuales debido a diferencias en el tamaño traqueideo). Los árboles que crecen en climas sin estaciones bien expresadas, como la selva tropical, no harán anillos anuales en absoluto.
Las maderas duras son producidas por angiospermas y contienen elementos de vasos y traqueides (figura\(\PageIndex{10}\)). Las maderas blandas son producidas por coníferas (en el filo gimnospermo Coniferophyta) y contienen solo traqueides (Figura\(\PageIndex{11}\)). Estos términos son incorrectos en cierta medida, sin embargo, porque las maderas duras no siempre son más densas que las maderas blandas. Los rayos de xilema tienden a ocupar un mayor volumen en maderas duras en relación con las maderas blandas. Adicionalmente, la disposición de las celdas aparece más desordenada en maderas duras debido al gran tamaño de los elementos de los vasos. Finalmente, las maderas blandas contienen conductos de resina (Figura\(\PageIndex{11}\)), los cuales contienen una sustancia espesa (resina) importante en defensa y respuesta a lesiones.
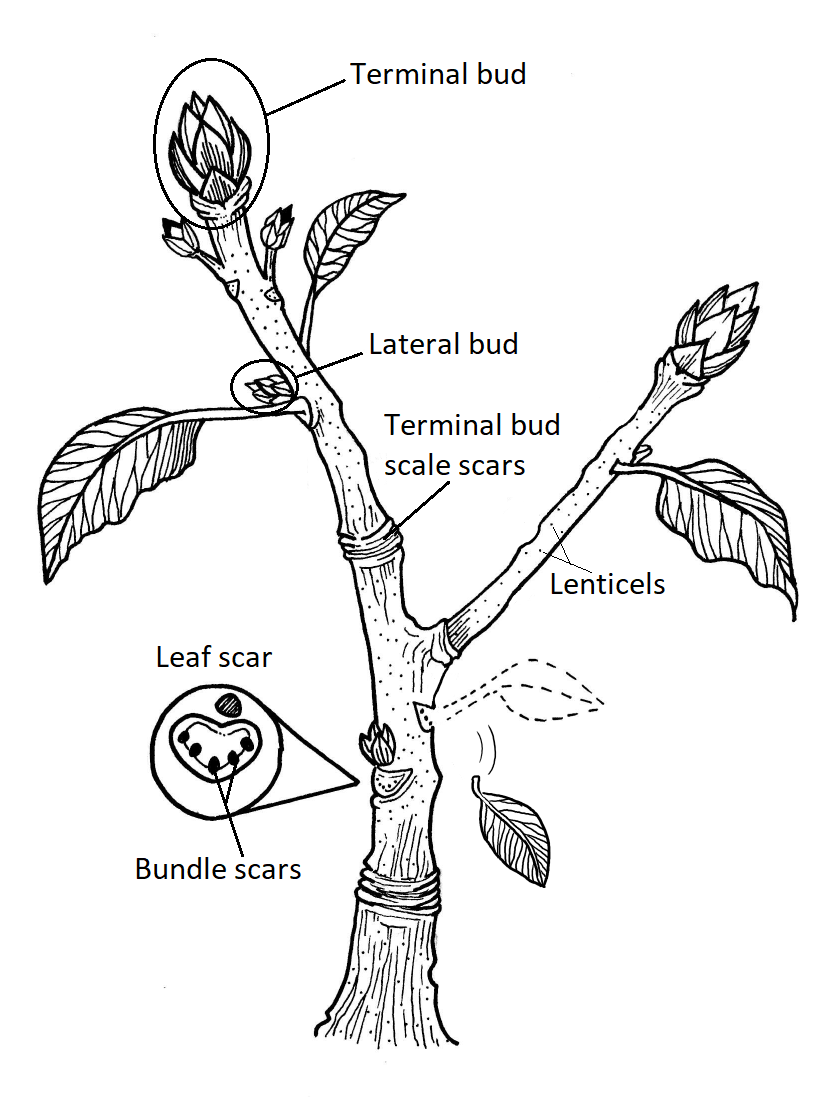
Ramitas de invierno
Los árboles y arbustos caducifolios invernales en regiones templadas se vuelven latentes en invierno. Las ramitas de estas especies tienen las características externas básicas de un tallo (yemas axilares, nódulos, etc.), pero se modifican para facilitar la latencia en el invierno y la reanudación del crecimiento en primavera. Al final de una ramita invernal se encuentra la yema terminal, que contiene un ápice de brote rodeado de estructuras protectoras llamadas escamas de cogollos. Cuando el brote terminal reanuda el crecimiento, las escamas de los cogollos se caen y dejan marcas llamadas cicatrices en escamas terminales. Estos forman un anillo alrededor de la ramita, marcando el invierno de cada año. Los cogollos laterales son similares en estructura a los cogollos terminales, pero se encuentran en cada nodo. Justo debajo de los cogollos laterales se encuentran cicatrices foliares, donde antes estaban adheridas las hojas. Dentro de las cicatrices foliares se encuentran cicatrices de haz, marcando rastros foliares (consistentes en haces vasculares) que se movían del tallo a la hoja (Figura\(\PageIndex{12}\)).
Atribuciones
Comisariada y autoría de Melissa Ha de las siguientes fuentes:
- De 5.4: La raíz de la introducción a la botánica de Alexey Shipunov (dominio público)
- De 30.2 Stems from Biology 2e por OpenStax (licenciado CC-BY)