
3.3: Características únicas de las células procariotas
- Page ID
- 54964

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Explicar las características distintivas de las células procariotas
- Describir las morfologías celulares comunes y las disposiciones celulares típicas de las células procariotas y explicar cómo las células mantienen su morfología
- Describir las estructuras internas y externas de las células procariotas en términos de su estructura física, estructura química y función
- Comparar las características distintivas de las células bacterianas y arqueales
La teoría celular afirma que la célula es la unidad fundamental de la vida. Sin embargo, las células varían significativamente en tamaño, forma, estructura y función. En el nivel de construcción más simple, todas las celdas poseen algunos componentes fundamentales. Estos incluyen citoplasma (una sustancia similar a un gel compuesta por agua y químicos disueltos necesarios para el crecimiento), que está contenido dentro de una membrana plasmática (también llamada membrana celular o membrana citoplásmica); uno o más cromosomas, que contienen los planos genéticos de la célula; y ribosomas, orgánulos utilizados para la producción de proteínas.
Más allá de estos componentes básicos, las células pueden variar mucho entre organismos, e incluso dentro del mismo organismo multicelular. Las dos categorías más grandes de células, las células procariotas y las células eucariotas, se definen por diferencias importantes en varias estructuras celulares. Las células procariotas carecen de un núcleo rodeado por una membrana nuclear compleja y generalmente tienen un solo cromosoma circular localizado en un nucleoide. Las células eucariotas tienen un núcleo rodeado por una membrana nuclear compleja que contiene múltiples cromosomas en forma de varilla. 1
Todas las células vegetales y animales son eucariotas. Algunos microorganismos están compuestos por células procariotas, mientras que otros están compuestos por células eucariotas. Los microorganismos procariotas se clasifican dentro de los dominios Archaea y Bacteria, mientras que los organismos eucariotas se clasifican dentro del dominio Eukarya.
Las estructuras dentro de una célula son análogas a los órganos dentro de un cuerpo humano, con estructuras únicas adecuadas para funciones específicas. Algunas de las estructuras que se encuentran en las células procariotas son similares a las que se encuentran en algunas células eucariotas; otras son exclusivas de procariotas. Aunque hay algunas excepciones, las células eucariotas tienden a ser más grandes que las células procariotas. El tamaño comparativamente mayor de las células eucariotas dicta la necesidad de compartimentar diversos procesos químicos dentro de diferentes áreas de la célula, utilizando orgánulos complejos unidos a la membrana. En contraste, las células procariotas generalmente carecen de orgánulos unidos a membrana; sin embargo, a menudo contienen inclusiones que compartimentalizan su citoplasma. La Figura\(\PageIndex{1}\) ilustra estructuras típicamente asociadas con células procariotas. Estas estructuras se describen con más detalle en la siguiente sección.
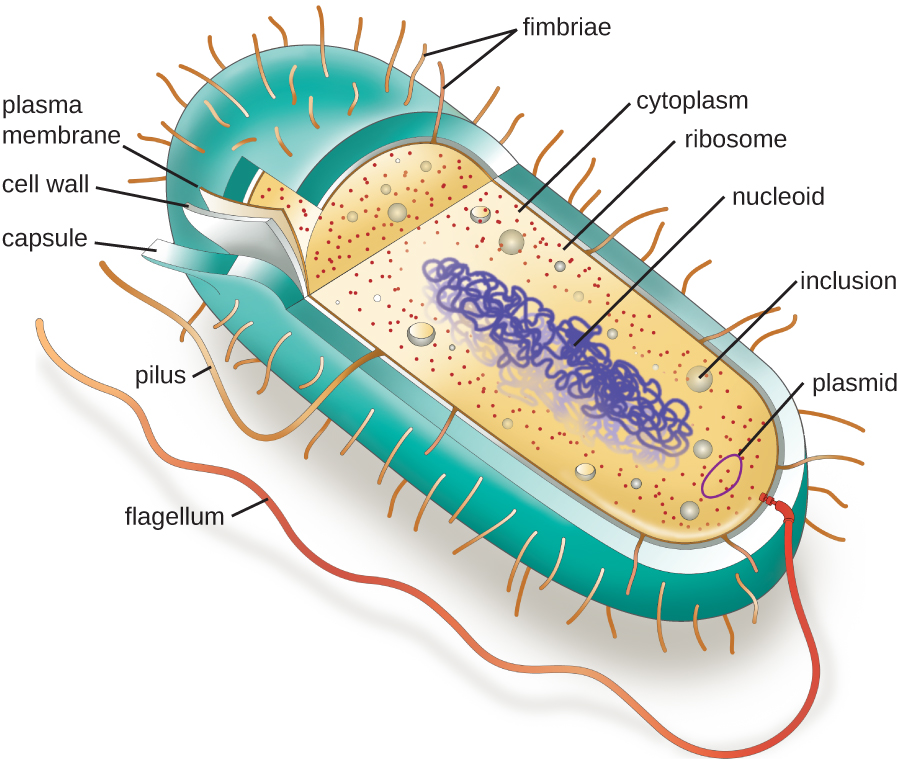
Morfologías y arreglos celulares comunes
Las células individuales de un organismo procariota particular son típicamente similares en forma o morfología celular. Aunque se han identificado miles de organismos procariotas, solo un puñado de morfologías celulares se ven comúnmente microscópicamente. La figura\(\PageIndex{2}\) nombra e ilustra las morfologías celulares que se encuentran comúnmente en células procariotas. Además de la forma celular, las células procariotas de la misma especie pueden agruparse en ciertos arreglos distintivos dependiendo del plano de división celular. Algunas disposiciones comunes se muestran en la Figura\(\PageIndex{3}\).
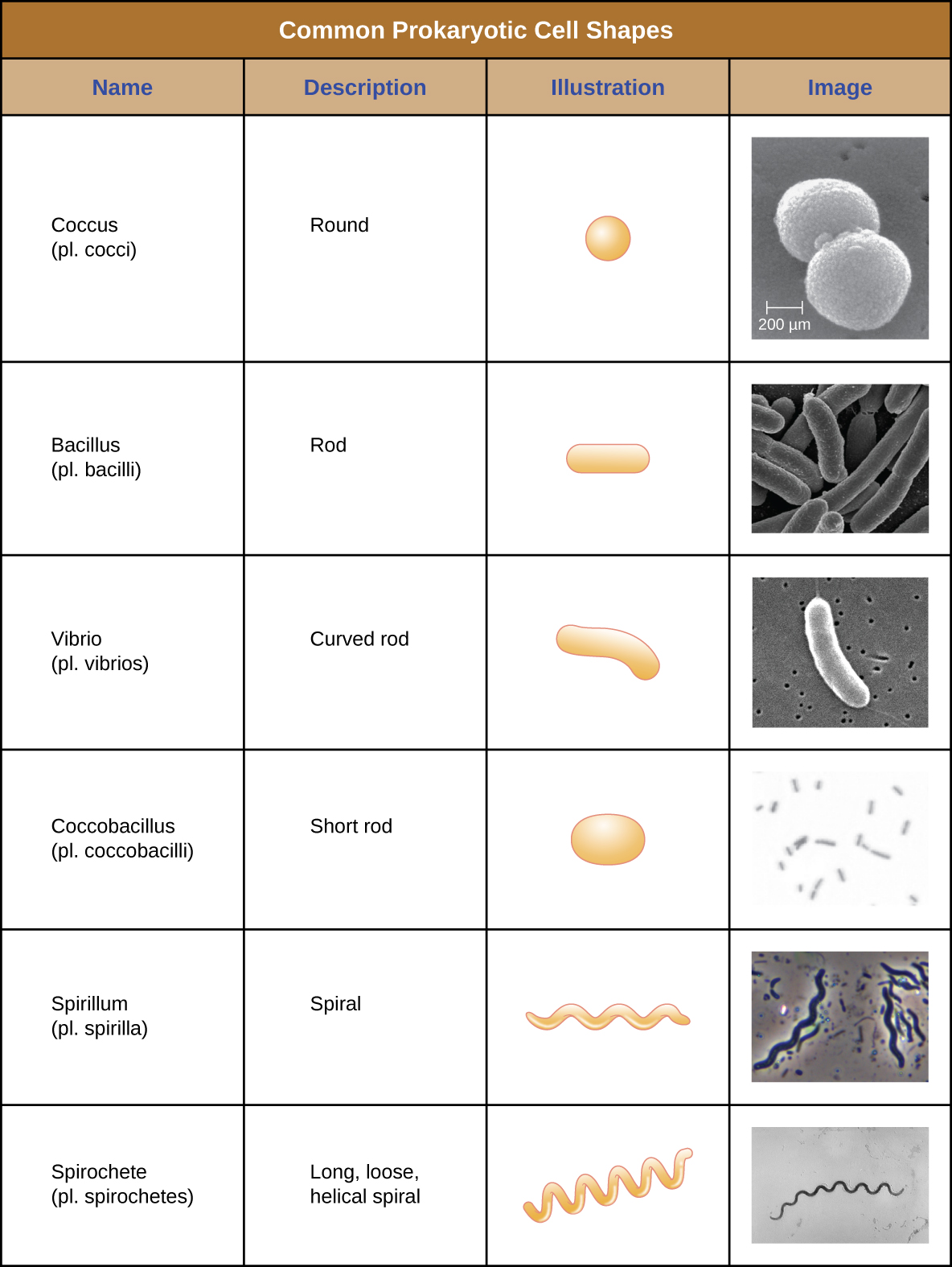
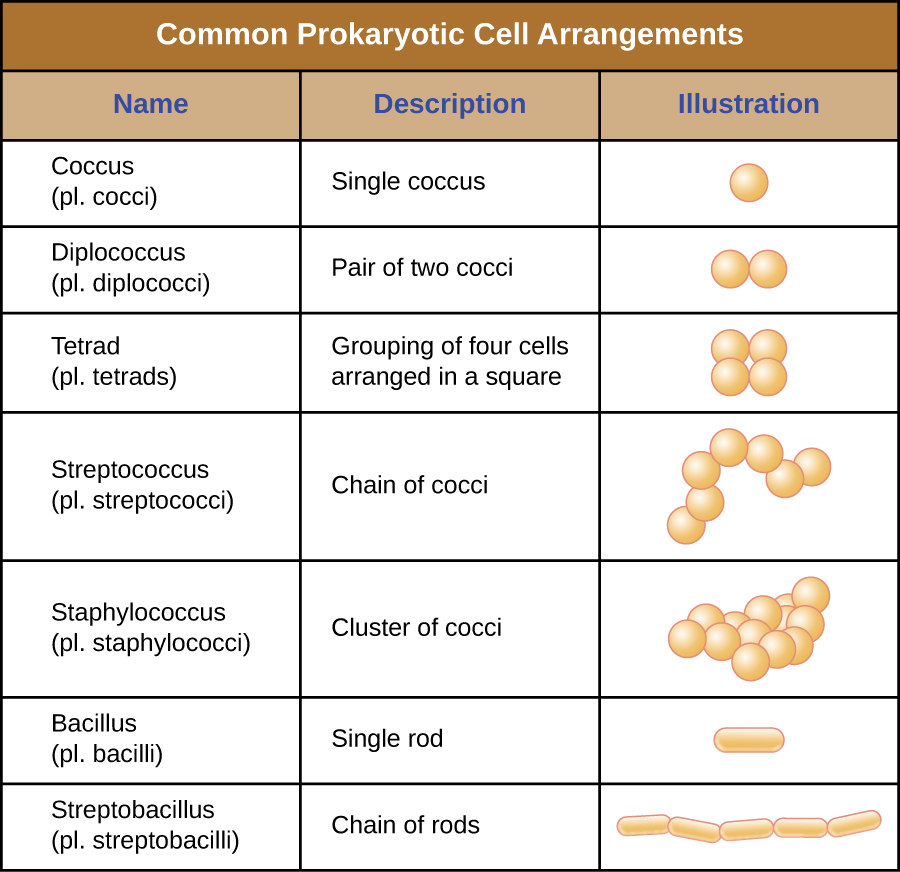
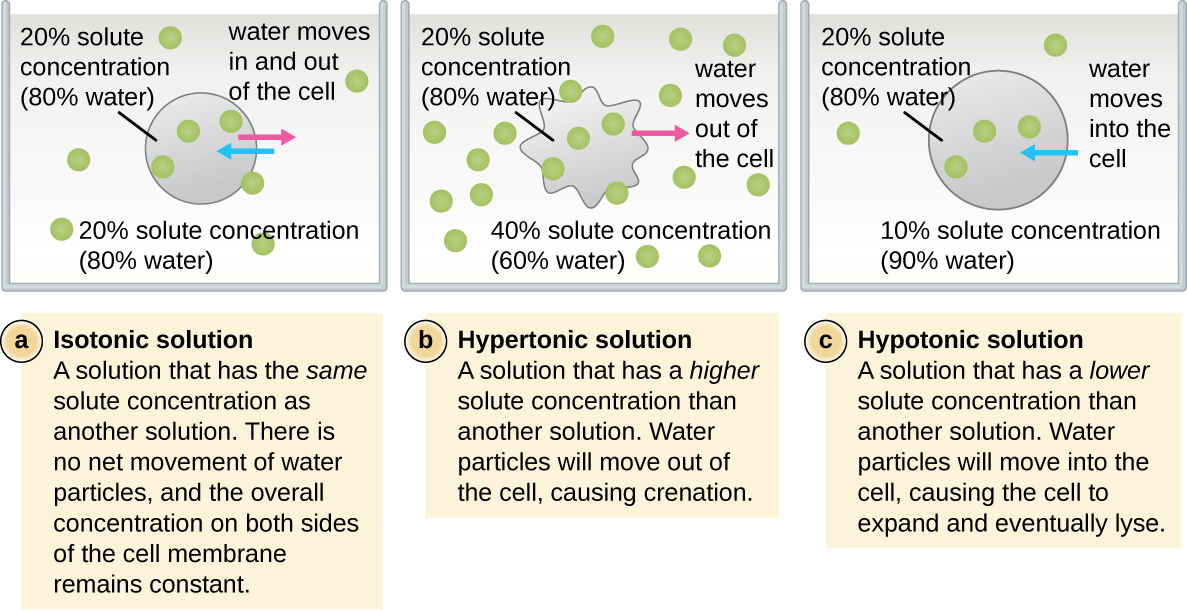
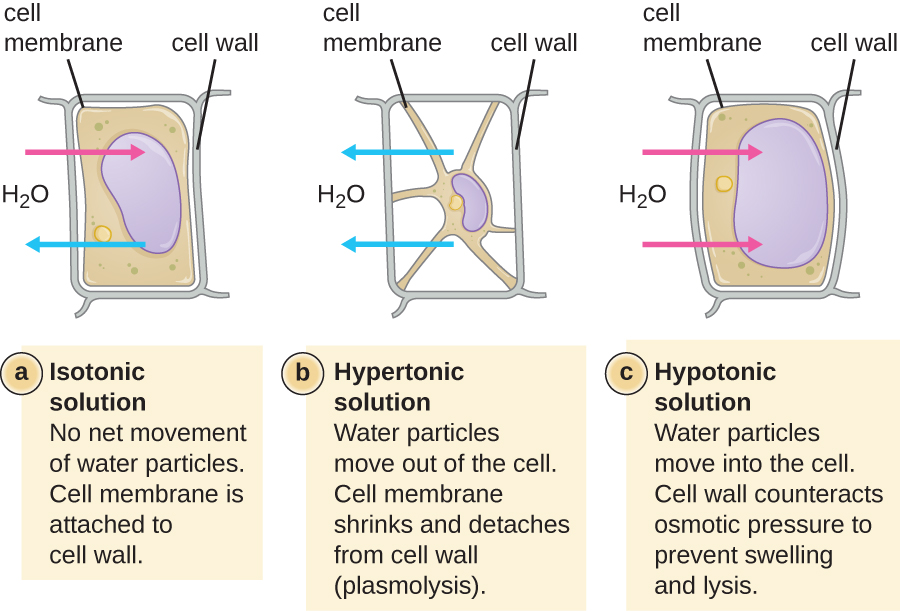
En la mayoría de las células procariotas, la morfología es mantenida por la pared celular en combinación con elementos citoesqueléticos. La pared celular es una estructura que se encuentra en la mayoría de los procariotas y algunos eucariotas; envuelve la membrana celular, protegiendo a la célula de los cambios en la presión osmótica (Figura\(\PageIndex{4}\)). La presión osmótica ocurre debido a diferencias en la concentración de solutos en lados opuestos de una membrana semipermeable. El agua es capaz de pasar a través de una membrana semipermeable, pero los solutos (moléculas disueltas como sales, azúcares y otros compuestos) no pueden. Cuando la concentración de solutos es mayor en un lado de la membrana, el agua se difunde a través de la membrana desde el lado con la concentración más baja (más agua) hacia el lado con la concentración más alta (menos agua) hasta que las concentraciones en ambos lados se vuelven iguales. Esta difusión del agua se llama ósmosis, y puede causar una presión osmótica extrema sobre una célula cuando cambia su ambiente externo.
El ambiente externo de una célula puede describirse como un medio isotónico, hipertónico o hipotónico. En un medio isotónico, las concentraciones de soluto dentro y fuera de la célula son aproximadamente iguales, por lo que no hay movimiento neto de agua a través de la membrana celular. En un medio hipertónico, la concentración de soluto fuera de la célula excede la del interior de la célula, por lo que el agua se difunde fuera de la célula y hacia el medio externo. En un medio hipotónico, la concentración de soluto dentro de la célula supera a la del exterior de la célula, por lo que el agua se moverá por ósmosis hacia la célula. Esto hace que la célula se hinche y potencialmente lise, o estalle.
El grado en que una célula en particular es capaz de soportar cambios en la presión osmótica se llama tonicidad. Las células que tienen una pared celular son más capaces de soportar cambios sutiles en la presión osmótica y mantener su forma. En ambientes hipertónicos, las células que carecen de pared celular pueden deshidratarse, causando almenación, o marchitamiento de la célula; la membrana plasmática se contrae y aparece festoneada o entallada (Figura\(\PageIndex{4}\)). Por el contrario, las células que poseen una pared celular experimentan plasmólisis en lugar de almenación. En la plasmólisis, la membrana plasmática se contrae y se desprende de la pared celular, y hay una disminución en el volumen interior, pero la pared celular permanece intacta, permitiendo así que la célula mantenga cierta forma e integridad por un periodo de tiempo (Figura\(\PageIndex{5}\)). Asimismo, las células que carecen de una pared celular son más propensas a la lisis en ambientes hipotónicos. La presencia de una pared celular permite que la célula mantenga su forma e integridad durante más tiempo antes de lisar (Figura\(\PageIndex{5}\)).
Ejercicio\(\PageIndex{1}\)
- Explicar la diferencia entre la morfología celular y el arreglo.
- ¿Qué ventajas proporcionan las paredes celulares a las células procariotas?
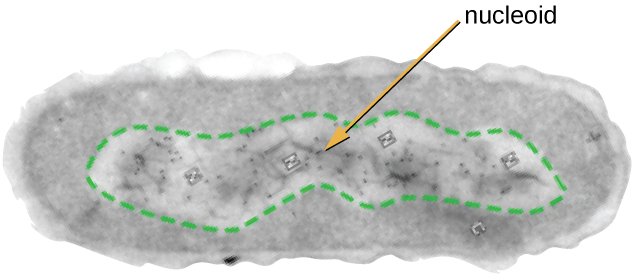
El Nucleoide
Toda la vida celular tiene un genoma de ADN organizado en uno o más cromosomas. Los cromosomas procariotas son típicamente circulares, haploides (desapareados) y no unidos por una membrana nuclear compleja. El ADN procariota y las proteínas asociadas al ADN se concentran dentro de la región nucleoide de la célula (Figura\(\PageIndex{6}\)). En general, el ADN procariota interactúa con proteínas asociadas a nucleósidos (NAP) que ayudan en la organización y empaquetamiento del cromosoma. En las bacterias, los NAP funcionan de manera similar a las histonas, que son las proteínas organizadoras del ADN que se encuentran en las células eucariotas. En las arqueas, el nucleoide está organizado por NAP o proteínas organizadoras de ADN similares a histonas.
Plásmidos
Las células procariotas también pueden contener ADN extracromosómico, o ADN que no es parte del cromosoma. Este ADN extracromosómico se encuentra en plásmidos, que son moléculas de ADN pequeñas, circulares y bicatenarias. Las células que tienen plásmidos suelen tener cientos de ellas dentro de una sola célula. Los plásmidos se encuentran más comúnmente en bacterias; sin embargo, los plásmidos se han encontrado en arqueas y organismos eucariotas. Los plásmidos a menudo portan genes que confieren rasgos ventajosos como la resistencia a antibióticos; por lo tanto, son importantes para la supervivencia del organismo. Discutiremos plásmidos con más detalle en Mecanismos de Genética Microbiana.
Ribosomas
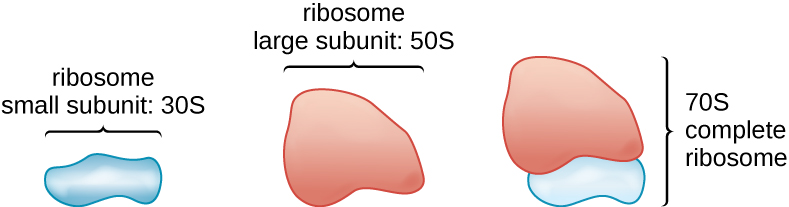
Toda la vida celular sintetiza proteínas, y los organismos en los tres dominios de la vida poseen ribosomas, estructuras responsables de la síntesis de proteínas. Sin embargo, los ribosomas en cada uno de los tres dominios son estructuralmente diferentes. Los ribosomas, ellos mismos, se construyen a partir de proteínas, junto con ARN ribosómico (ARNr). Los ribosomas procariotas se encuentran en el citoplasma. Se les llama ribosomas 70S porque tienen un tamaño de 70S (Figura\(\PageIndex{7}\)), mientras que los ribosomas citoplásmicos eucariotas tienen un tamaño de 80S. (La S significa unidad Svedberg, una medida de sedimentación en una ultracentrífuga, que se basa en el tamaño, la forma y las cualidades superficiales de la estructura que se analiza). Aunque son del mismo tamaño, los ribosomas bacterianos y arqueales tienen diferentes proteínas y moléculas de ARNr, y las versiones arqueales son más similares a sus contrapartes eucariotas que a las que se encuentran en las bacterias.
Inclusiones
Como organismos unicelulares que viven en ambientes inestables, algunas células procariotas tienen la capacidad de almacenar el exceso de nutrientes dentro de estructuras citoplásmicas llamadas inclusiones. El almacenamiento de nutrientes en forma polimerizada es ventajoso porque reduce la acumulación de presión osmótica que se produce a medida que una célula acumula solutos. Varios tipos de inclusiones almacenan glucógeno y almidones, los cuales contienen carbono al que las células pueden acceder para obtener energía. Los gránulos de volutina, también llamados gránulos metacromáticos por sus características de tinción, son inclusiones que almacenan fosfato inorgánico polimerizado que puede ser utilizado en el metabolismo y ayudar en la formación de biopelículas. Los microbios que se sabe que contienen gránulos de volutina incluyen la archaea Methanosarcina, la bacteria Corynebacterium diphtheriae y la alga eucariota unicelular Chlamydomonas. Los gránulos de azufre, otro tipo de inclusión, se encuentran en bacterias de azufre del género Thiobacillus; estos gránulos almacenan azufre elemental, que las bacterias utilizan para el metabolismo.
Ocasionalmente, ciertos tipos de inclusiones están rodeadas por una monocapa de fosfolípidos incrustada con proteína. El polihidroxibutirato (PHB), que puede ser producido por especies de Bacillus y Pseudomonas, es un ejemplo de una inclusión que muestra este tipo de estructura monocapa. Industrialmente, el PHB también se ha utilizado como fuente de polímeros biodegradables para bioplásticos. En la Figura se muestran varios tipos diferentes de inclusiones\(\PageIndex{8}\).

Algunas células procariotas tienen otro tipo de inclusiones que sirven para fines distintos al almacenamiento de nutrientes. Por ejemplo, algunas células procariotas producen vacuolas de gas, acumulaciones de pequeñas vesículas de gas revestidas de proteínas. Estas vacuolas de gas permiten que las células procariotas que las sintetizan alteren su flotabilidad para que puedan ajustar su ubicación en la columna de agua. Las bacterias magnetotácticas, como Magnetospirillum magnetotacticum, contienen magnetosomas, que son inclusiones de óxido de hierro magnético o sulfuro de hierro rodeadas por una capa lipídica. Estos permiten que las células se alineen a lo largo de un campo magnético, lo que ayuda a su movimiento (Figura\(\PageIndex{8}\)). Cianobacterias como Anabaena cylindrica y bacterias como Halothiobacillus neapolitanus producen inclusiones de carboxisomas. Los carboxisomas están compuestos por conchas externas de miles de subunidades proteicas. Su interior está lleno de ribulosa-1,5-bisfosfato carboxilasa/oxigenasa (RubiSco) y anhidrasa carbónica. Ambos compuestos se utilizan para el metabolismo del carbono. Algunas células procariotas también poseen carboxisomas que secuestran enzimas funcionalmente relacionadas en una ubicación. Estas estructuras se consideran proto-orgánulos porque compartimentan compuestos importantes o reacciones químicas, al igual que muchos orgánulos eucariotas.
Endosporas
Las células bacterianas generalmente se observan como células vegetativas, pero algunos géneros de bacterias tienen la capacidad de formar endosporas, estructuras que esencialmente protegen el genoma bacteriano en estado latente cuando las condiciones ambientales son desfavorables. Las endosporas (que no deben confundirse con las esporas reproductivas formadas por hongos) permiten que algunas células bacterianas sobrevivan largos periodos sin alimentos ni agua, así como la exposición a productos químicos, temperaturas extremas e incluso radiación. En el cuadro\(\PageIndex{1}\) se comparan las características de las células vegetativas y endosporas.
| Células Vegetativas | Endosporas |
|---|---|
| Sensible a temperaturas extremas y radiación | Resistente a temperaturas extremas y radiación |
| Gram-positivos | No absorber la mancha de Gram, solo las manchas especiales de endosporas (ver Tinción de especímenes microscópicos) |
| Contenido normal de agua y actividad enzimática | Deshidratado; sin actividad metabólica |
| Capaz de crecimiento activo y metabolismo | Inactivo; sin crecimiento ni actividad metabólica |
El proceso por el cual las células vegetativas se transforman en endosporas se llama esporulación, y generalmente comienza cuando los nutrientes se agotan o las condiciones ambientales se vuelven desfavorables de otro modo (Figura\(\PageIndex{9}\)). El proceso comienza con la formación de un tabique en la célula bacteriana vegetativa. El tabique divide la célula asimétricamente, separando una forespora de ADN de la célula madre. La forespora, que formará el núcleo de la endospora, es esencialmente una copia de los cromosomas de la célula, y está separada de la célula madre por una segunda membrana. Una corteza se forma gradualmente alrededor de la forespora colocando capas de calcio y ácido dipicolínico entre las membranas. Luego se forma una cubierta de esporas proteicas alrededor de la corteza mientras el ADN de la célula madre se desintegra. La maduración adicional de la endospora ocurre con la formación de un exosporio más externo. La endospora se libera tras la desintegración de la célula madre, completando la esporulación.
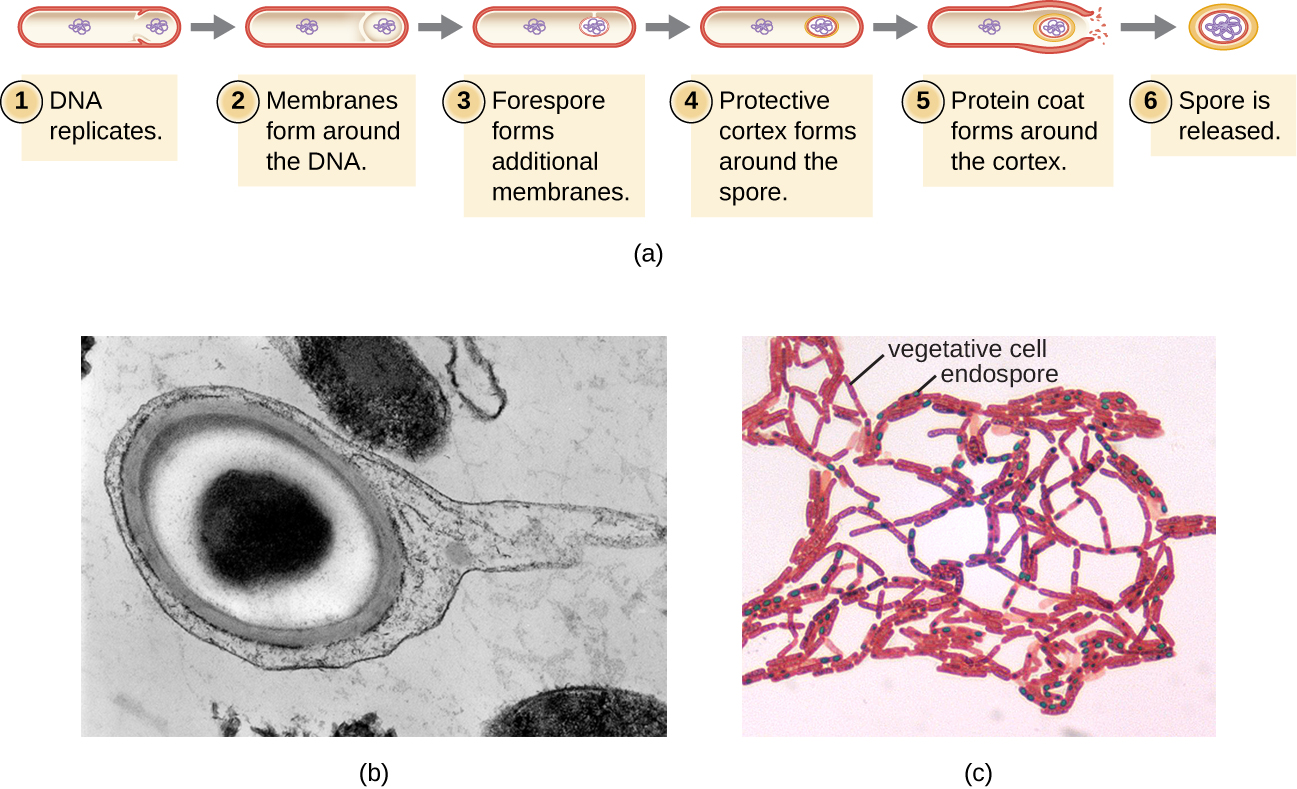
Se ha demostrado que las endosporas de ciertas especies persisten en estado latente por periodos prolongados de tiempo, hasta miles de años. 2 Sin embargo, cuando las condiciones de vida mejoran, las endosporas se someten a germinación, reingresando a un estado vegetativo. Después de la germinación, la célula vuelve a ser metabólicamente activa y es capaz de llevar a cabo todas sus funciones normales, incluyendo el crecimiento y la división celular.
No todas las bacterias tienen la capacidad de formar endosporas; sin embargo, hay una serie de bacterias grampositivas formadoras de endosporas clínicamente significativas de los géneros Bacillus y Clostridium. Estos incluyen B. anthracis, el agente causante del ántrax, que produce endosporas capaces de sobrevivir por muchas décadas 3; C. tetani (causa tétanos); C. difficile (causa colitis pseudomembranosa); C. perfringens (causa gangrena gaseosa); y C. botulinum (causa botulismo). Patógenos como estos son particularmente difíciles de combatir porque sus endosporas son muy difíciles de matar. Métodos especiales de esterilización para bacterias formadoras de endosporas se discuten en Control de Crecimiento Microbiano.
Ejercicio\(\PageIndex{2}\)
- ¿Qué es una inclusión?
- ¿Cuál es la función de una endospora?
Membrana Plasma
Las estructuras que encierran el citoplasma y las estructuras internas de la célula se conocen colectivamente como la envoltura celular. En las células procariotas, las estructuras de la envoltura celular varían dependiendo del tipo de célula y organismo. La mayoría (pero no todas) las células procariotas tienen una pared celular, pero la composición de esta pared celular varía. Todas las células (procariotas y eucariotas) tienen una membrana plasmática (también llamada membrana citoplásmica o membrana celular) que exhibe permeabilidad selectiva, permitiendo que algunas moléculas entren o salgan de la célula al tiempo que restringe el paso de otras.
La estructura de la membrana plasmática a menudo se describe en términos del modelo de mosaico de fluidos, que se refiere a la capacidad de los componentes de la membrana para moverse fluidamente dentro del plano de la membrana, así como la composición similar a un mosaico de los componentes, que incluyen una diversa matriz de componentes lipídicos y proteicos ( Figura\(\PageIndex{10}\)). La estructura de la membrana plasmática de la mayoría de los tipos de células bacterianas y eucariotas es una bicapa compuesta principalmente por fosfolípidos formados con enlaces éster y proteínas. Estos fosfolípidos y proteínas tienen la capacidad de moverse lateralmente dentro del plano de las membranas así como entre las dos capas de fosfolípidos.

Las membranas arqueales son fundamentalmente diferentes de las membranas bacterianas y eucariotas en algunas formas significativas. Primero, los fosfolípidos de membrana arqueal se forman con enlaces éter, en contraste con los enlaces éster que se encuentran en las membranas celulares bacterianas o eucariotas. Segundo, los fosfolípidos arqueales tienen cadenas ramificadas, mientras que los de las células bacterianas y eucariotas son de cadena lineal. Finalmente, aunque algunas membranas arqueales pueden estar formadas por bicapas como las que se encuentran en bacterias y eucariotas, otras membranas plasmáticas arqueales son monocapas lipídicas.
Las proteínas en la superficie celular son importantes para una variedad de funciones, incluyendo la comunicación célula a célula, y la detección de condiciones ambientales y factores de virulencia patógena. Las proteínas de membrana y los fosfolípidos pueden tener carbohidratos (azúcares) asociados a ellas y se denominan glicoproteínas o glicolípidos, respectivamente. Estos complejos de glicoproteínas y glicolípidos se extienden desde la superficie de la célula, permitiendo que la célula interactúe con el entorno externo (Figura\(\PageIndex{10}\)). Las glicoproteínas y glicolípidos en la membrana plasmática pueden variar considerablemente en composición química entre arqueas, bacterias y eucariotas, lo que permite a los científicos utilizarlas para caracterizar especies únicas.
Las membranas plasmáticas de diferentes tipos de células también contienen fosfolípidos únicos, que contienen ácidos grasos. Como se describe en Uso de la bioquímica para identificar microorganismos, los perfiles de análisis de ácidos grasos derivados de fosfolípidos (PLFA) pueden usarse para identificar tipos únicos de células basadas en diferencias en los ácidos grasos. Las arqueas, bacterias y eucariotas tienen cada una un perfil PFLA único.
Mecanismos de transporte de membrana
Una de las funciones más importantes de la membrana plasmática es controlar el transporte de moléculas dentro y fuera de la célula. Las condiciones internas deben mantenerse dentro de un cierto rango a pesar de cualquier cambio en el entorno externo. El transporte de sustancias a través de la membrana plasmática permite que las células lo hagan.
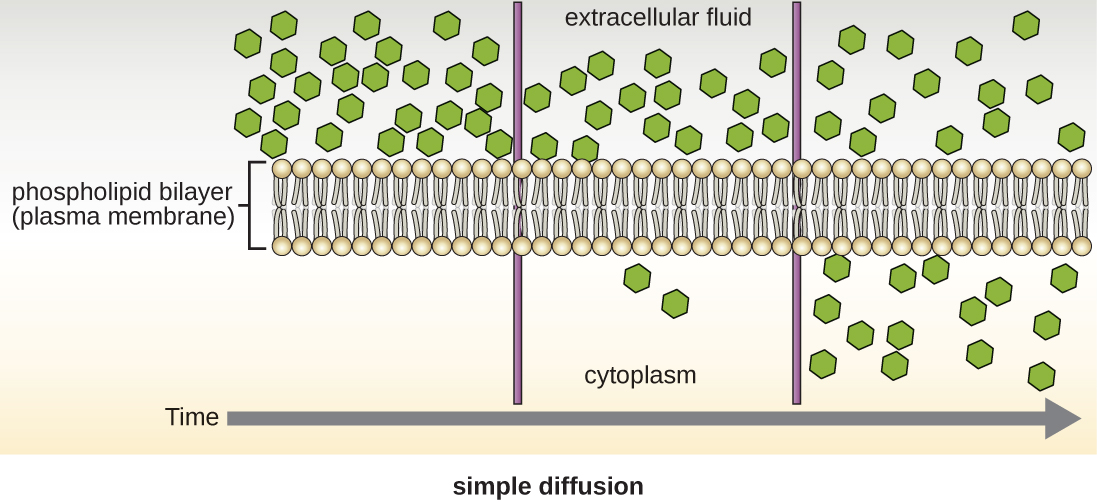
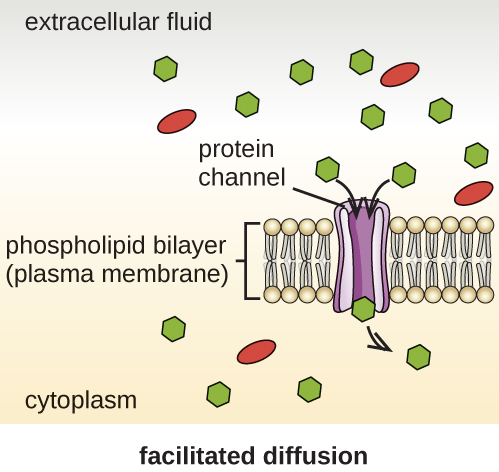
Las células utilizan varios modos de transporte a través de la membrana plasmática. Por ejemplo, las moléculas que pasan de una concentración mayor a una concentración menor con el gradiente de concentración son transportadas por difusión simple, también conocida como transporte pasivo (Figura\(\PageIndex{11}\)). Algunas moléculas pequeñas, como el dióxido de carbono, pueden cruzar la bicapa de membrana directamente por simple difusión. Sin embargo, las moléculas cargadas, así como las moléculas grandes, necesitan la ayuda de portadores o canales en la membrana. Estas estructuras transportan moléculas a través de la membrana, un proceso conocido como difusión facilitada (Figura\(\PageIndex{12}\)).
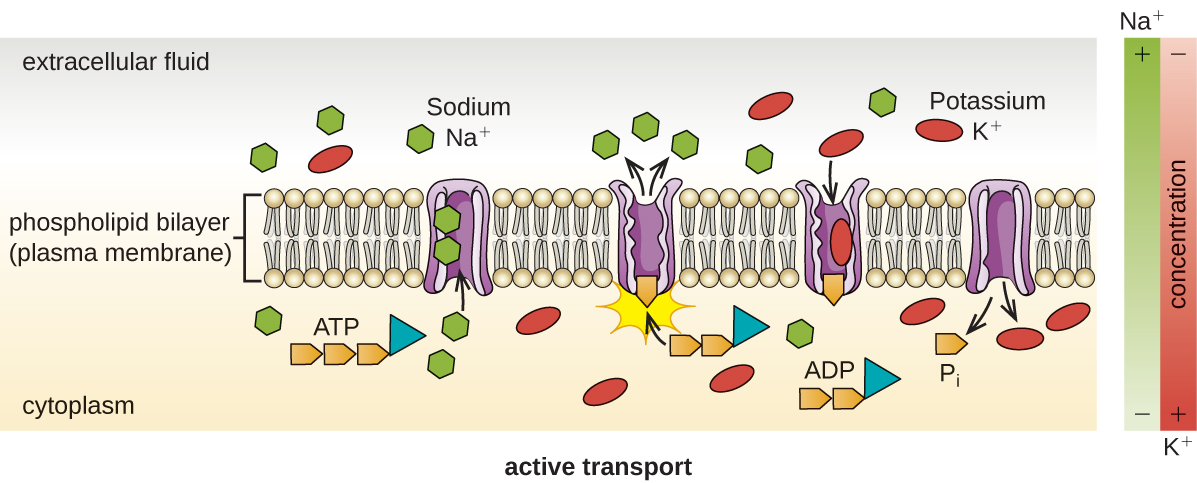
El transporte activo ocurre cuando las células mueven moléculas a través de su membrana contra gradientes de concentración (Figura\(\PageIndex{13}\)). Una diferencia importante entre el transporte pasivo y el activo es que el transporte activo requiere trifosfato de adenosina (ATP) u otras formas de energía para mover las moléculas “cuesta arriba”. Por lo tanto, las estructuras de transporte activo a menudo se llaman “bombas”.
La translocación grupal también transporta sustancias al interior de las células bacterianas. En este caso, a medida que una molécula se mueve hacia una célula frente a su gradiente de concentración, se modifica químicamente para que no requiera transporte contra un gradiente de concentración desfavorable. Un ejemplo común de esto es el sistema bacteriano fosfotransferasa, una serie de portadores que fosforila (es decir, añade iones fosfato a) glucosa u otros azúcares al entrar en las células. Dado que se requiere la fosforilación de azúcares durante las primeras etapas del metabolismo del azúcar, se considera que el sistema de fosfotransferasa es un sistema de energía neutra.
Estructuras de membrana fotosintéticas
Algunas células procariotas, a saber, cianobacterias y bacterias fotosintéticas, tienen estructuras de membrana que les permiten realizar fotosíntesis. Estas estructuras consisten en un infolding de la membrana plasmática que encierra pigmentos fotosintéticos como clorofilas verdes y bacterioclorofilas. En las cianobacterias, estas estructuras de membrana se llaman tilacoides; en las bacterias fotosintéticas, se les llama cromatóforos, laminillas o clorosomas.
Pared celular
La función principal de la pared celular es proteger la célula de las duras condiciones en el ambiente exterior. Cuando están presentes, hay notables similitudes y diferencias entre las paredes celulares de arqueas, bacterias y eucariotas.
El componente principal de las paredes celulares bacterianas se llama peptidoglicano (o mureína); solo se encuentra en bacterias. Estructuralmente, el peptidoglicano se asemeja a una capa de malla o tela (Figura\(\PageIndex{14}\)). Cada capa está compuesta por largas cadenas de moléculas alternas de N-acetilglucosamina (NAG) y ácido N-acetilmurámico (NAM). La estructura de las cadenas largas tiene una resistencia a la tracción bidimensional significativa debido a la formación de puentes peptídicos que conectan NAG y NAM dentro de cada capa de peptidoglicano. En las bacterias gramnegativas, las cadenas de tetrapéptidos que se extienden desde cada unidad NAM están directamente reticuladas, mientras que en las bacterias grampositivas, estas cadenas de tetrapéptidos están unidas por puentes cruzados de pentaglicina. Las subunidades de peptidoglicano se hacen dentro de la célula bacteriana y luego se exportan y ensamblan en capas, dando a la célula su forma.
Dado que el peptidoglicano es exclusivo de las bacterias, muchos antibióticos están diseñados para interferir con la síntesis de peptidoglicanos, debilitando la pared celular y haciendo que las células bacterianas sean más susceptibles a los efectos de la presión osmótica (ver Mecanismos de Medicamentos Antibacterianos). Además, ciertas células del sistema inmune humano son capaces de “reconocer” patógenos bacterianos al detectar peptidoglicano en la superficie de una célula bacteriana; estas células luego engullen y destruyen la célula bacteriana, utilizando enzimas como la lisozima, que descompone y digiere el peptidoglicano en sus paredes celulares (ver Reconocimiento de Patógenos y Fagocitosis).
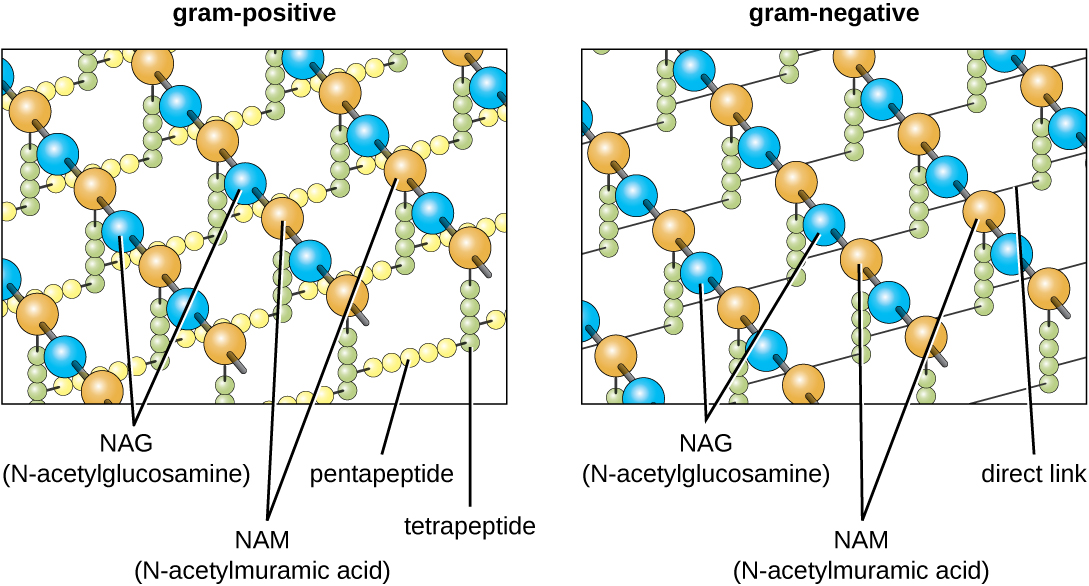
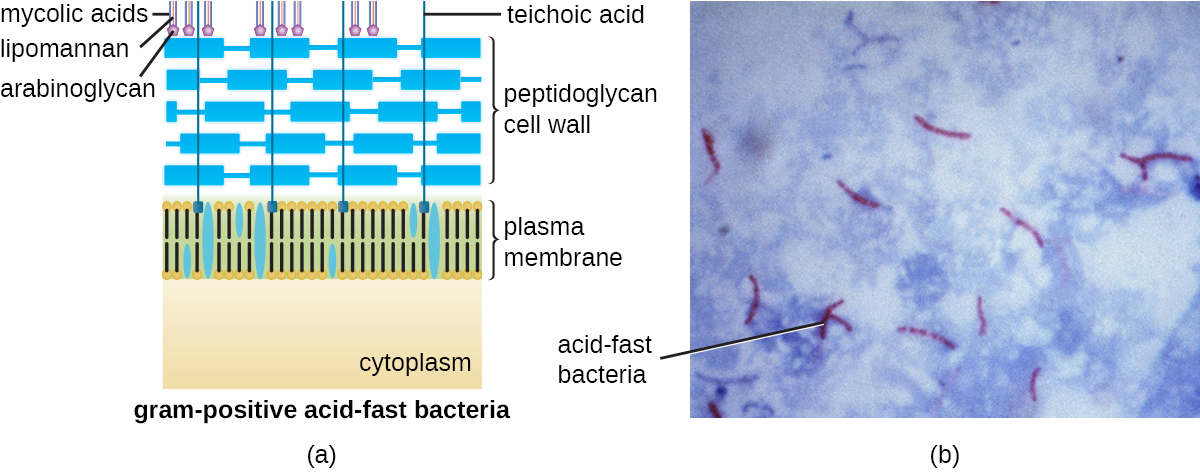
El protocolo de tinción de Gram (ver Tinción de Muestras Microscópicas) se utiliza para diferenciar dos tipos comunes de estructuras de pared celular (Figura\(\PageIndex{15}\)). Las células grampositivas tienen una pared celular que consiste en muchas capas de peptidoglicano con un grosor total de 30—100 nm. Estas capas de peptidoglicano están comúnmente incrustadas con ácidos teicoicos (TA), cadenas de carbohidratos que se extienden a través y más allá de la capa de peptidoglicano. 4 Se cree que el TA estabiliza el peptidoglicano aumentando su rigidez. El TA también juega un papel en la capacidad de bacterias grampositivas patógenas como Streptococcus para unirse a ciertas proteínas en la superficie de las células hospedadoras, potenciando su capacidad para causar infección. Además de peptidoglicano y TA, las bacterias de la familia Mycobacteriaceae tienen una capa externa de ácidos micólicos cerosos en su pared celular; como se describe en Tinción de especímenes microscópicos, estas bacterias son referidas como ácido-resistentes, ya que las manchas ácido-resistentes deben usarse para penetrar el micólico capa ácida para fines de microscopía (Figura\(\PageIndex{16}\)).

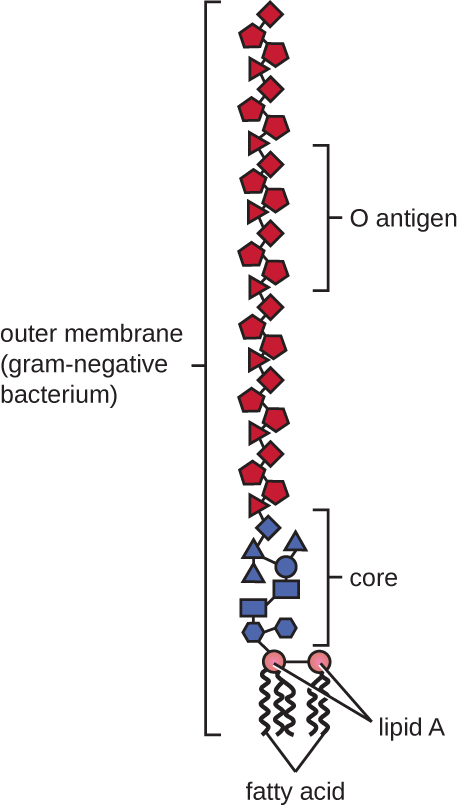
Las células gramnegativas tienen una capa mucho más delgada de peptidoglicano (no más de aproximadamente 4 nm de espesor 6) que las células grampositivas, y la estructura general de su envoltura celular es más compleja. En las células gramnegativas, una matriz similar a un gel ocupa el espacio periplásmico entre la pared celular y la membrana plasmática, y existe una segunda bicapa lipídica llamada membrana externa, que es externa a la capa de peptidoglicano (Figura\(\PageIndex{15}\)). Esta membrana externa está unida al peptidoglicano por la lipoproteína mureína. La valva externa de la membrana externa contiene la molécula lipopolisacárido (LPS), que funciona como endotoxina en infecciones que involucran bacterias gramnegativas, contribuyendo a síntomas como fiebre, hemorragia y choque séptico. Cada molécula de LPS está compuesta por Lípido A, un polisacárido central y una cadena lateral O que está compuesta por moléculas similares a azúcar que comprenden la cara externa del LPS (Figura\(\PageIndex{17}\)). La composición de la cadena lateral O varía entre diferentes especies y cepas de bacterias. Partes de la cadena lateral O llamadas antígenos pueden detectarse mediante pruebas serológicas o inmunológicas para identificar cepas patógenas específicas como Escherichia coli O157:H7, una cepa mortal de bacterias que causa diarrea sanguinolenta e insuficiencia renal.
La estructura arqueal de la pared celular difiere de la de las bacterias en varias formas significativas. Primero, las paredes celulares arqueales no contienen peptidoglicano; en cambio, contienen un polímero similar llamado pseudopeptidoglicano (pseudomureína) en el que el NAM es reemplazado por una subunidad diferente. Otras arqueas pueden tener una capa de glicoproteínas o polisacáridos que sirve como pared celular en lugar de pseudopeptidoglicano. Por último, como es el caso de algunas especies bacterianas, hay algunas arqueas que parecen carecer de paredes celulares por completo.
Glicocálices y capas S
Aunque la mayoría de las células procariotas tienen paredes celulares, algunas pueden tener estructuras adicionales de envoltura celular exteriores a la pared celular, como glicocálices y capas S. Un glicocáliz es una capa de azúcar, de la cual existen dos tipos importantes: cápsulas y capas de limo. Una cápsula es una capa organizada ubicada fuera de la pared celular y generalmente compuesta por polisacáridos o proteínas (Figura\(\PageIndex{18}\)). Una capa de limo es una capa menos organizada que solo se adhiere flojamente a la pared celular y se puede lavar más fácilmente. Las capas de limo pueden estar compuestas de polisacáridos, glicoproteínas o glicolípidos.
Los glicocálices permiten que las células se adhieran a las superficies, ayudando en la formación de biopelículas (colonias de microbios que se forman en capas sobre superficies). En la naturaleza, la mayoría de los microbios viven en comunidades mixtas dentro de biopelículas, en parte porque la biopelícula les brinda cierto nivel de protección. Las biopelículas generalmente retienen el agua como una esponja, evitando la desecación. También protegen a las células de la depredación y obstaculizan la acción de antibióticos y desinfectantes. Todas estas propiedades son ventajosas para los microbios que viven en una biopelícula, pero presentan desafíos en un entorno clínico, donde el objetivo suele ser eliminar microbios.

La capacidad de producir una cápsula puede contribuir a la patogenicidad de un microbio (capacidad de causar enfermedades) porque la cápsula puede dificultar que las células fagocíticas (como los glóbulos blancos) engulle y maten al microorganismo. Streptococcus pneumoniae, por ejemplo, produce una cápsula que es bien conocida por ayudar en la patogenicidad de esta bacteria. Como se explica en Tinción de Muestras Microscópicas, las cápsulas son difíciles de teñir para microscopía; típicamente se utilizan técnicas de tinción negativa.
Una capa S es otro tipo de estructura de envoltura celular; está compuesta por una mezcla de proteínas estructurales y glicoproteínas. En las bacterias, las capas S se encuentran fuera de la pared celular, pero en algunas arqueas, la capa S sirve como pared celular. La función exacta de las capas S no se entiende del todo, y son difíciles de estudiar; pero la evidencia disponible sugiere que pueden desempeñar una variedad de funciones en diferentes células procariotas, como ayudar a la célula a soportar la presión osmótica y, para ciertos patógenos, interactuar con el sistema inmune del huésped sistema.
Enfoque Clínico: Parte 3
Después de diagnosticar a Bárbara con neumonía, la AP le escribe una receta para amoxicilina, un tipo de derivado de penicilina comúnmente prescrito. Más de una semana después, a pesar de tomar el curso completo como se le indica, Bárbara todavía se siente débil y no está completamente recuperada, aunque todavía es capaz de superar sus actividades diarias. Regresa al centro de salud para una visita de seguimiento.
Muchos tipos de bacterias, hongos y virus pueden causar neumonía. La amoxicilina se dirige al peptidoglicano de las paredes celulares bacterianas. Dado que la amoxicilina no ha resuelto los síntomas de Barbara, la AP concluye que el agente causal probablemente carece de peptidoglicano, lo que significa que el patógeno podría ser un virus, un hongo o una bacteria que carece de peptidoglicano. Otra posibilidad es que el patógeno sea una bacteria que contiene peptidoglicano pero ha desarrollado resistencia a la amoxicilina.
Ejercicio\(\PageIndex{3}\)
- ¿Cómo puede la AP identificar definitivamente la causa de la neumonía de Bárbara?
- ¿Qué forma de tratamiento debe prescribir la AP, dado que la amoxicilina era ineficaz?
Apéndices Filamentosos
Muchas células bacterianas tienen apéndices proteicos incrustados dentro de sus envolturas celulares que se extienden hacia afuera, permitiendo la interacción con el ambiente. Estos apéndices pueden adherirse a otras superficies, transferir ADN o proporcionar movimiento. Los apéndices filamentosos incluyen fimbrias, pilos y flagelos.
Fimbriae y Pili
Fimbriae y pili son estructuralmente similares y, debido a que la diferenciación entre los dos es problemática, estos términos a menudo se usan indistintamente. 7 8 El término fimbrias se refiere comúnmente a proteínas cortas parecidas a cerdas que se proyectan de la superficie celular por cientos. Las fimbrias permiten que una celda se adhiera a superficies y a otras celdas. Para las bacterias patógenas, la adherencia a las células hospedadoras es importante para la colonización, infectividad y virulencia. La adherencia a las superficies también es importante en la formación de biopelículas.
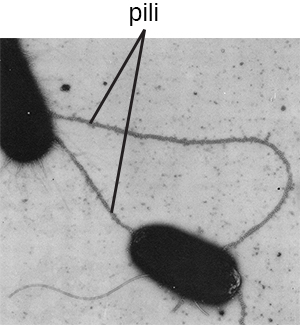
El término pili (singular: pilus) comúnmente se refiere a apéndices proteicos más largos y menos numerosos que ayudan en la unión a las superficies (Figura\(\PageIndex{19}\)). Un tipo específico de pilus, llamado el pilus F o pilus sexual, es importante en la transferencia de ADN entre células bacterianas, lo que ocurre entre miembros de una misma generación cuando dos células transfieren o intercambian físicamente partes de sus respectivos genomas (ver Cómo los procariotas asexuales logran genética Diversidad).
Antes de que se entendiera bien la estructura y función de los diversos componentes de la envoltura celular bacteriana, los científicos ya estaban utilizando las características de la envoltura celular para clasificar las bacterias. En 1933, Rebecca Lancefield propuso un método para serotipar diversas cepas β-hemolíticas de especies de Streptococcus mediante un ensayo de aglutinación, una técnica que utiliza la aglutinación de bacterias para detectar antígenos específicos de la superficie celular. Al hacerlo, Lancefield descubrió que un grupo de S. pyogenes, que se encuentra en el Grupo A, estaba asociado con una variedad de enfermedades humanas. Determinó que diversas cepas del estreptococo del Grupo A podrían distinguirse entre sí en base a variaciones en proteínas específicas de la superficie celular a las que llamó proteínas M.
Hoy en día, se han identificado más de 80 cepas diferentes de estreptocócicos del Grupo A a partir de proteínas M. Diversas cepas de estreptococos del grupo A están asociadas con una amplia variedad de infecciones humanas, incluyendo faringitis estreptocócica (faringitis estreptocócica), impétigo, síndrome de choque tóxico, escarlatina, fiebre reumática y fascitis necrotizante. La proteína M es un factor de virulencia importante para el estreptococo del Grupo A, ayudando a estas cepas a evadir el sistema inmunológico. Los cambios en las proteínas M parecen alterar la infectividad de una cepa particular del estreptocócico del Grupo A.
Flagelos
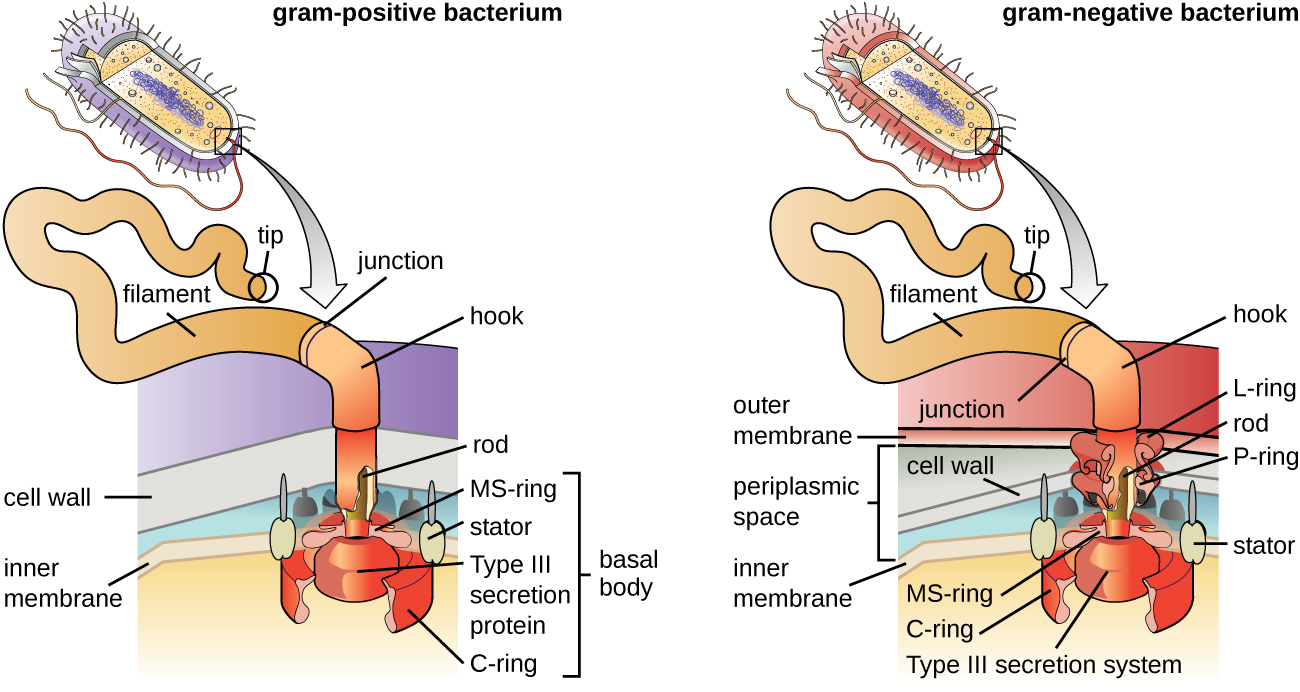
Los flagelos son estructuras utilizadas por las células para moverse en ambientes acuosos. Los flagelos bacterianos actúan como hélices. Son filamentos espirales rígidos compuestos por subunidades de proteína flagelina que se extienden hacia afuera desde la célula y giran en solución. El cuerpo basal es el motor para el flagelo y está incrustado en la membrana plasmática (Figura\(\PageIndex{20}\)). Una región de gancho conecta el cuerpo basal con el filamento. Las bacterias grampositivas y gramnegativas tienen diferentes configuraciones basales corporales debido a diferencias en la estructura de la pared celular.
Diferentes tipos de bacterias móviles exhiben diferentes disposiciones de flagelos (Figura\(\PageIndex{21}\)). Se dice que una bacteria con un flagelo singular, típicamente localizada en un extremo de la célula (polar), tiene un flagelo monotríco. Un ejemplo de un patógeno bacteriano flagelado monotricamente es Vibrio cholerae, la bacteria gramnegativa que causa el cólera. Las células con flagelos anfitricos tienen un flagelo o mechones de flagelos en cada extremo. Un ejemplo es Spirillum minor, la causa de la fiebre espírita (asiática) por mordedura de rata o sodoku. Las células con flagelos lofotricos tienen un penacho en un extremo de la célula. El bacilo gramnegativo Pseudomonas aeruginosa, un patógeno oportunista conocido por causar muchas infecciones, incluyendo “oído de nadador” e infecciones por quemaduras, tiene flagelos lofotricos. Los flagelos que cubren toda la superficie de una célula bacteriana se denominan flagelos peritricos. La bacteria gramnegativa E. coli muestra un arreglo peririco de flagelos.
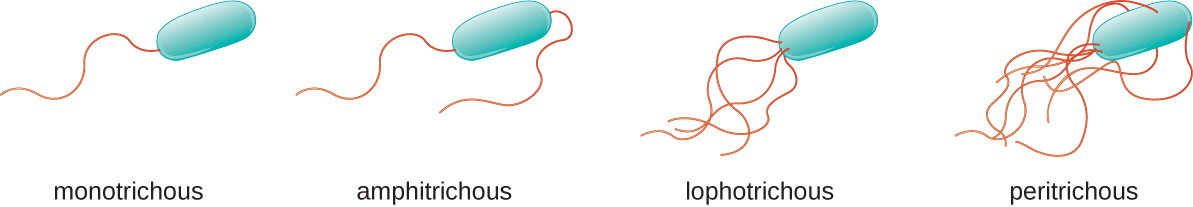
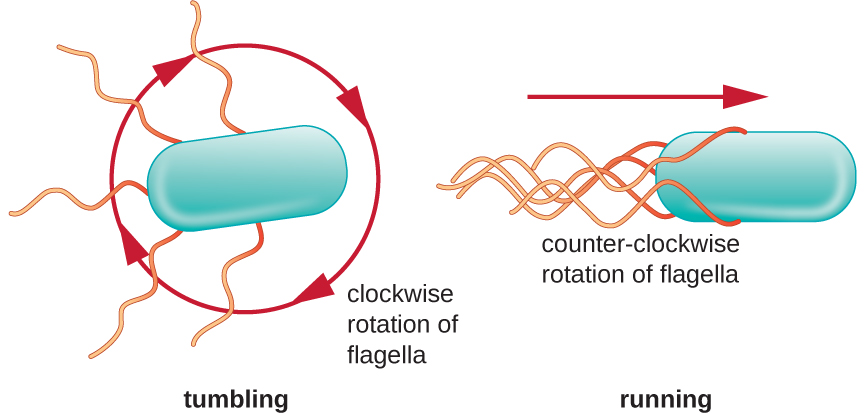
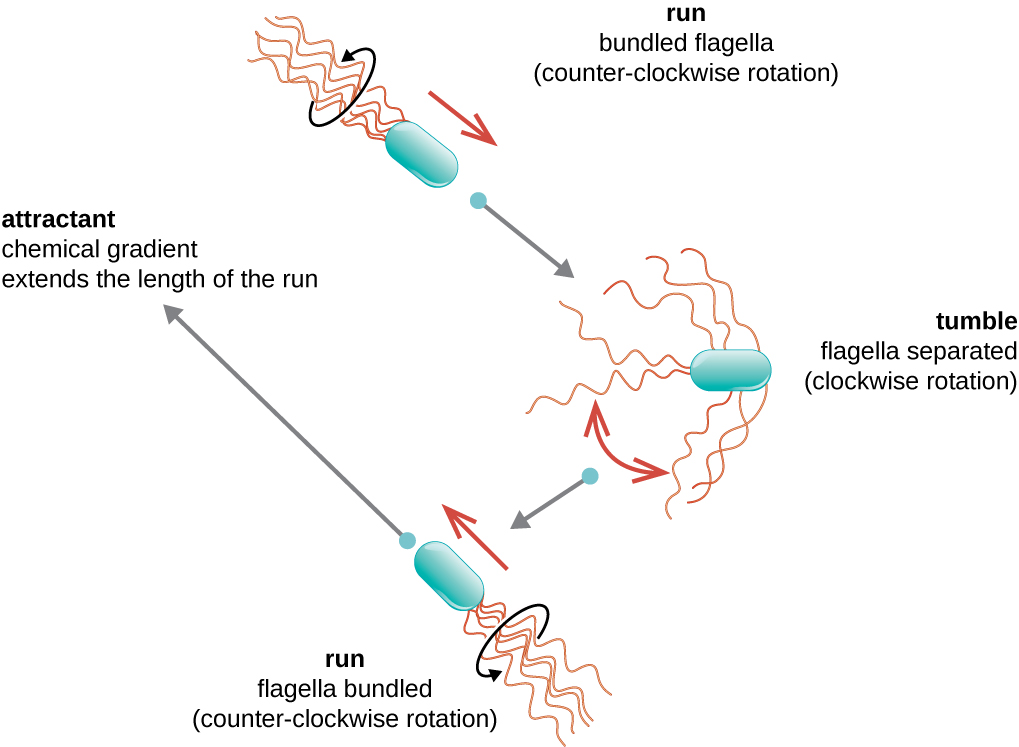
El movimiento direccional depende de la configuración de los flagelos. Las bacterias pueden moverse en respuesta a una variedad de señales ambientales, incluyendo luz (fototaxis), campos magnéticos (magnetotaxis) usando magnetosomas y, más comúnmente, gradientes químicos (quimiotaxis). El movimiento intencionado hacia un atrayente químico, como una fuente de alimento, o lejos de un repelente, como un químico venenoso, se logra aumentando la longitud de las corridas y disminuyendo la longitud de los volteos. Al correr, los flagelos giran en sentido contrario a las agujas del reloj, permitiendo que la célula bacteriana avance. En una bacteria peritrichosa, los flagelos están todos agrupados de una manera muy aerodinámica (Figura\(\PageIndex{22}\)), permitiendo un movimiento eficiente. Al caer, los flagelos se extienden mientras giran en el sentido de las agujas del reloj, creando un movimiento de bucle y evitando un movimiento significativo hacia adelante pero reorientando la célula hacia la dirección del atrayente. Cuando existe un atrayente, aún ocurren corridas y volteos; sin embargo, la longitud de las corridas es más larga, mientras que la longitud de los volteos se reduce, permitiendo el movimiento general hacia la mayor concentración del atrayente. Cuando no existe gradiente químico, las longitudes de corridas y volteos son más iguales, y el movimiento general es más aleatorio (Figura\(\PageIndex{23}\)).
Ejercicio\(\PageIndex{4}\)
- ¿Qué es la capa de peptidoglicano y en qué se diferencia entre bacterias grampositivas y gramnegativas?
- Comparar y contrastar flagelos monotricos, anfitricos, lofotricos y peritricos.
Resumen
- Las células procariotas difieren de las células eucariotas en que su material genético está contenido en un nucleoide en lugar de en un núcleo unido a la membrana. Además, las células procariotas generalmente carecen de orgánulos unidos a membrana.
- Las células procariotas de la misma especie suelen compartir una morfología celular y una disposición celular similares.
- La mayoría de las células procariotas tienen una pared celular que ayuda al organismo a mantener la morfología celular y lo protege contra los cambios en la presión osmótica.
- Fuera del nucleoide, las células procariotas pueden contener ADN extracromosómico en plásmidos.
- Los ribosomas procariotas que se encuentran en el citoplasma tienen un tamaño de 70S.
- Algunas células procariotas tienen inclusiones que almacenan nutrientes o químicos para otros usos.
- Algunas células procariotas son capaces de formar endosporas a través de la esporulación para sobrevivir en un estado latente cuando las condiciones son desfavorables. Las endosporas pueden germinar, transformándose de nuevo en células vegetativas cuando mejoran las condiciones.
- En las células procariotas, la envoltura celular incluye una membrana plasmática y generalmente una pared celular.
- Las membranas bacterianas están compuestas por fosfolípidos con proteínas integrales o periféricas. Los componentes ácidos grasos de estos fosfolípidos están unidos a éster y a menudo se utilizan para identificar tipos específicos de bacterias. Las proteínas cumplen una variedad de funciones, incluyendo el transporte, la comunicación célula a célula y la detección de las condiciones ambientales. Las membranas arqueales son distintas en que están compuestas de ácidos grasos que están unidos por éteres a fosfolípidos.
- Algunas moléculas pueden moverse a través de la membrana bacteriana por simple difusión, pero la mayoría de las moléculas grandes deben transportarse activamente a través de estructuras de membrana usando energía celular.
- Las paredes celulares procariotas pueden estar compuestas por peptidoglicano (bacteria) o pseudopeptidoglicano (arqueas).
- Las células bacterianas grampositivas se caracterizan por una capa gruesa de peptidoglicano, mientras que las células bacterianas gramnegativas se caracterizan por una fina capa de peptidoglicano rodeada por una membrana externa.
- Algunas células procariotas producen recubrimientos de glicocáliz, como cápsulas y capas de limo, que ayudan en la unión a las superficies y/o la evasión del sistema inmune del huésped.
- Algunas células procariotas tienen fimbrias o pili, apéndices filamentosos que ayudan en la fijación a las superficies. Los pili también se utilizan en la transferencia de material genético entre células.
- Algunas células procariotas utilizan uno o más flagelos para moverse a través del agua. Las bacterias peritróficas, que tienen numerosos flagelos, utilizan corridas y caídas para moverse a propósito en la dirección de un atrayente químico.
Notas al pie
- 1 Y.-H.M. Chan, W. F. Marshall. “Propiedades de Escalamiento del Tamaño de Celda y Organelo”. Organogénesis 6 núm. 2 (2010) :88—96.
- 2 F. Rothfuss, M Bender, R Conrad. “Supervivencia y Actividad de Bacterias en un Sedimento Lacustre Profundo y Envejecido (Lago Constanza). Ecología Microbiana 33 núm. 1 (1997) :69—77.
- 3 R. Sinclair et al. “Persistencia de Agentes Selectivos de Categoría A en el Medio Ambiente”. Microbiología Aplicada y Ambiental 74 núm. 3 (2008) :555—563.
- 4 T.J. Silhavy, D. Kahne, S. Walker. “La Envoltura Celular Bacteriana”. Cold Spring Harbor Perspectivas en Biología 2 núm. 5 (2010) :a000414.
- 5 B. Zuber et al. “Capa Granular en el Espacio Periplásmico de Bacterias Gram-Positivas y Estructuras Finas de Septos de Enterococcus gallinarum y Streptococcus gordonii Revelada por Crio-Microscopía Electrónica de Secciones Vítreas”. Revista de Bacteriología 188 núm. 18 (2006) :6652—6660
- 6 L. Gana, S. Chena, G.J. Jensena. “Organización Molecular del Peptidoglicano Gram-Negativo”. Actas de la Academia Nacional de Ciencias de los Estados Unidos de América 105 núm. 48 (2008) :18953—18957.
- 7 J.A. Garnetta et al. “Perspectivas estructurales sobre la biogénesis y la formación de biopelículas por el pilus común de Escherichia coli”. Actas de la Academia Nacional de Ciencias de los Estados Unidos de América 109 núm. 10 (2012) :3950—3955.
- 8 T. Proft, E.N. Baker. “Pili en Bacterias Gram-negativas y Gram-Positivas—Estructura, Ensamblaje y Su Papel en la Enfermedad”. Ciencias de la Vida Celular y Molecular 66 (2009) :613.