
3.4: Características únicas de las células eucariotas
- Page ID
- 54958

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Explicar las características distintivas de las células eucariotas
- Describir las estructuras internas y externas de las células procariotas en términos de su estructura física, estructura química y función
- Identificar y describir estructuras y orgánulos exclusivos de células eucariotas
- Comparar y contrastar estructuras similares encontradas en células procariotas y eucariotas
Los organismos eucariotas incluyen protozoos, algas, hongos, plantas y animales. Algunas células eucariotas son microorganismos unicelulares independientes, mientras que otras forman parte de organismos multicelulares. Las células de organismos eucariotas tienen varias características distintivas. Sobre todo, las células eucariotas se definen por la presencia de un núcleo rodeado por una membrana nuclear compleja. Además, las células eucariotas se caracterizan por la presencia de orgánulos unidos a membrana en el citoplasma. Organelos como las mitocondrias, el retículo endoplásmico (ER), el aparato de Golgi, los lisosomas y los peroxisomas se mantienen en su lugar por el citoesqueleto, una red interna que soporta el transporte de componentes intracelulares y ayuda a mantener la forma celular (Figura\(\PageIndex{1}\)). El genoma de las células eucariotas está empaquetado en múltiples cromosomas en forma de varilla, a diferencia del cromosoma único de forma circular que caracteriza a la mayoría de las células procariotas. La tabla\(\PageIndex{1}\) compara las características de las estructuras celulares eucariotas con las de bacterias y arqueas.

| Estructura celular | Procariotas | Eucariotas | |
|---|---|---|---|
| Bacterias | Archaea | ||
| Tamaño | ~0.5—1 μM | ~0.5—1 μM | ~5—20 μM |
| Relación de superficie a volumen | Alto | Alto | Bajo |
| Núcleo | No | No | Sí |
| Características del genoma |
|
|
|
| División celular | Fsión binaria | Fsión binaria | Mitosis, meiosis |
| Composición lipídica de membrana |
|
|
|
| Composición de la pared celular |
|
|
|
| Estructuras de motilidad | Flagelos espirales rígidos compuestos de flagelino | Flagelos espirales rígidos compuestos por flagelos arqueales | Flagelos flexibles y cilios compuestos por microtúbulos |
| Orgánulos unidos a membrana | No | No | Sí |
| Sistema Endomembrana | No | No | Sí (ER, Golgi, lisosomas) |
| Ribosomas | 70S | 70S |
|
Morfologías celulares
Las células eucariotas muestran una amplia variedad de morfologías celulares diferentes. Las formas posibles incluyen esferoides, ovoides, cuboidales, cilíndricas, planas, lenticulares, fusiformes, discoidales, de media luna, anulares estrelladas y poligonales (Figura\(\PageIndex{2}\)). Algunas células eucariotas son de forma irregular, y algunas son capaces de cambiar de forma. La forma de un tipo particular de célula eucariota puede estar influenciada por factores como su función primaria, la organización de su citoesqueleto, la viscosidad de su citoplasma, la rigidez de su membrana celular o pared celular (si tiene una) y la presión física ejercida sobre ella por el entorno circundante y/o celdas contiguas.
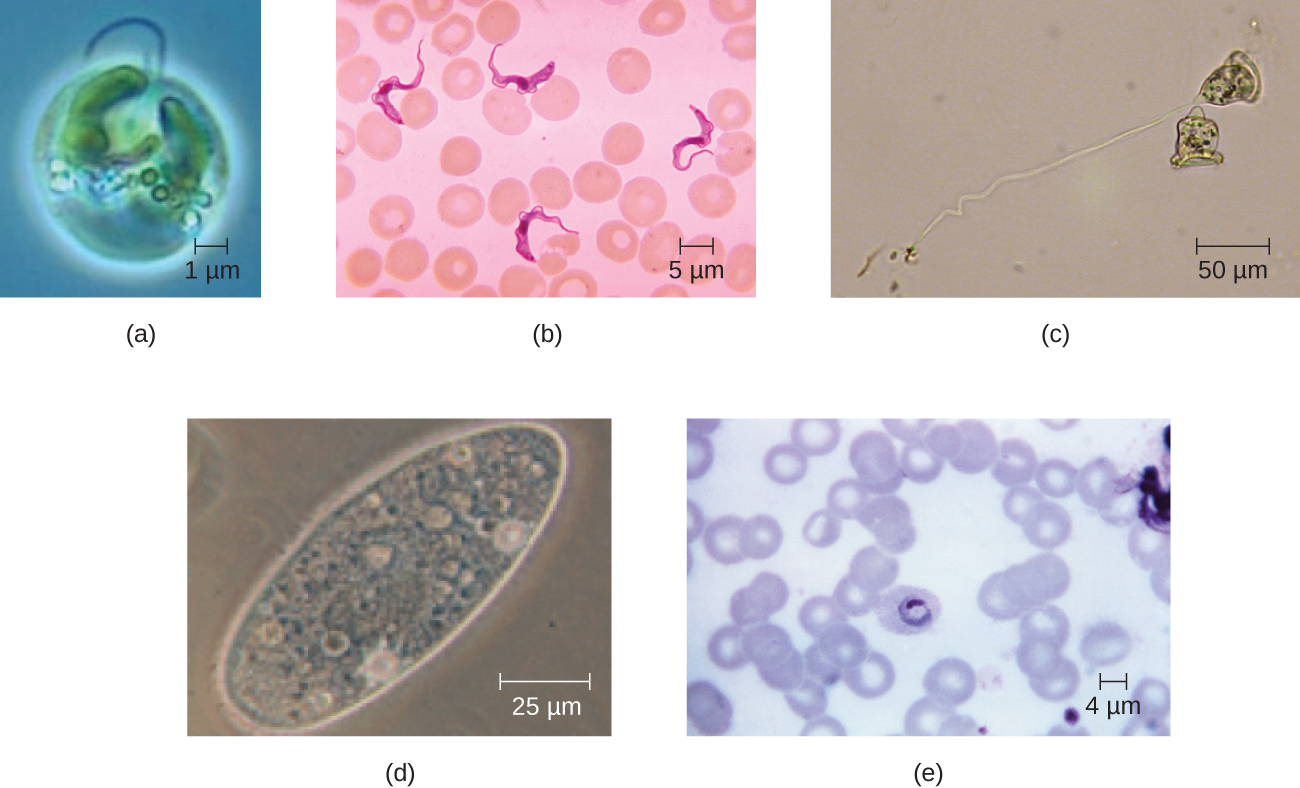
Ejercicio\(\PageIndex{1}\)
Identificar dos diferencias entre células eucariotas y procariotas.
Núcleo
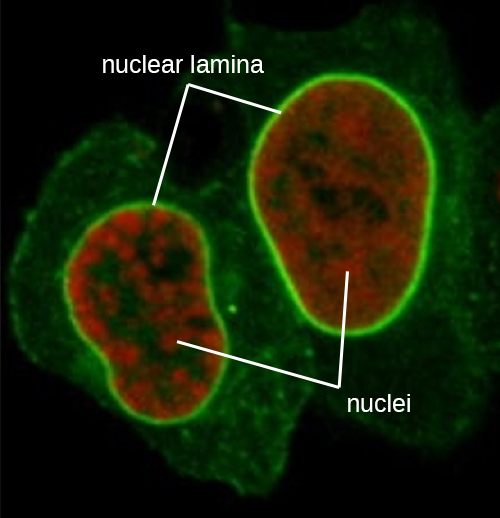
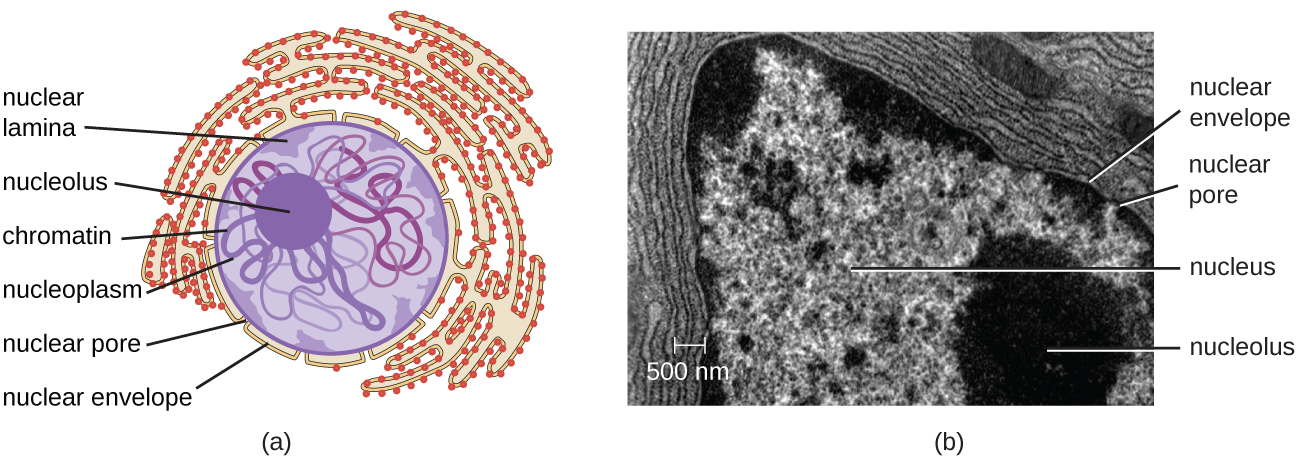
A diferencia de las células procariotas, en las que el ADN está contenido libremente en la región nucleoide, las células eucariotas poseen un núcleo, que está rodeado por una membrana nuclear compleja que alberga el genoma del ADN (Figura\(\PageIndex{3}\)). Al contener el ADN de la célula, el núcleo controla en última instancia todas las actividades de la célula y también cumple un papel esencial en la reproducción y la herencia. Las células eucariotas típicamente tienen su ADN organizado en múltiples cromosomas lineales. El ADN dentro del núcleo está altamente organizado y condensado para encajar dentro del núcleo, lo que se logra envolviendo el ADN alrededor de proteínas llamadas histonas.
Aunque la mayoría de las células eucariotas tienen un solo núcleo, existen excepciones. Por ejemplo, los protozoos del género Paramecium suelen tener dos núcleos completos: un núcleo pequeño que se utiliza para la reproducción (micronúcleos) y un núcleo grande que dirige el metabolismo celular (macronúcleo). Adicionalmente, algunos hongos forman transitoriamente células con dos núcleos, llamados células heterocarióticas, durante la reproducción sexual. Las células cuyos núcleos se dividen, pero cuyo citoplasma no, se llaman coenocitos.
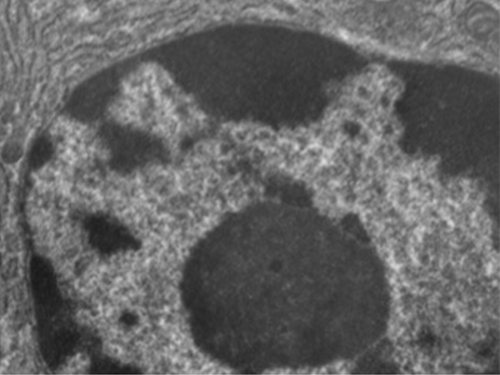
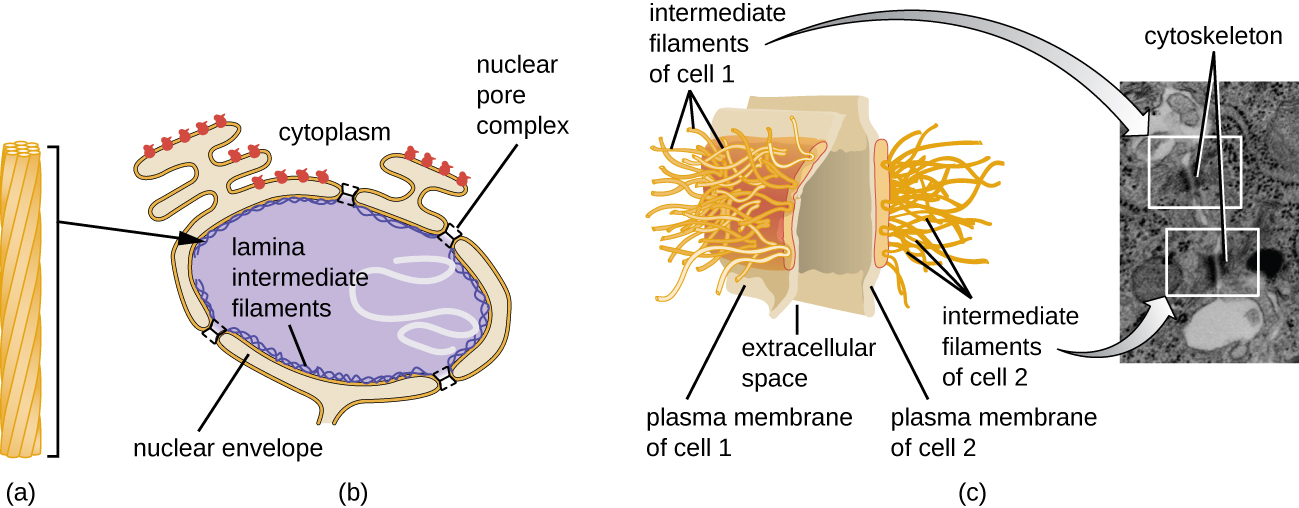
El núcleo está unido por una membrana nuclear compleja, a menudo llamada envoltura nuclear, que consiste en dos bicapas lipídicas distintas que son contiguas entre sí (Figura\(\PageIndex{4}\)). A pesar de estas conexiones entre las membranas interna y externa, cada membrana contiene lípidos y proteínas únicos en sus superficies interna y externa. La envoltura nuclear contiene poros nucleares, que son grandes complejos proteicos en forma de roseta que controlan el movimiento de los materiales dentro y fuera del núcleo. La forma general del núcleo está determinada por la lámina nuclear, una malla de filamentos intermedios que se encuentra justo dentro de las membranas de la envoltura nuclear. Fuera del núcleo, filamentos intermedios adicionales forman una malla más suelta y sirven para anclar el núcleo en posición dentro de la célula.
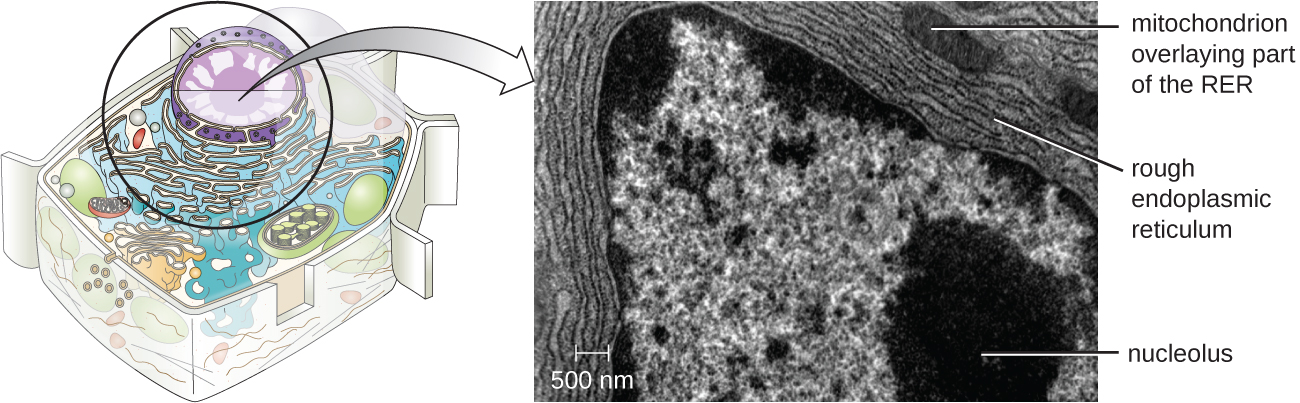
Nucleolo
El nucleolo es una región densa dentro del núcleo donde ocurre la biosíntesis de ARN ribosómico (ARNr). Además, el nucleolo es también el sitio donde comienza el ensamblaje de ribosomas. Los complejos preribosómicos se ensamblan a partir de ARNr y proteínas en el nucleolo; luego se transportan al citoplasma, donde se completa el ensamblaje del ribosoma (Figura\(\PageIndex{5}\)).
Ribosomas
Los ribosomas que se encuentran en orgánulos eucariotas como las mitocondrias o los cloroplastos tienen ribosomas 70S, el mismo tamaño que los ribosomas procariotas. Sin embargo, los ribosomas no asociados a orgánulos en células eucariotas son ribosomas 80S, compuestos por una subunidad pequeña 40S y una subunidad grande 60S. En términos de tamaño y composición, esto los hace distintos de los ribosomas de las células procariotas.
Los dos tipos de ribosomas eucariotas no asociados a orgánulos se definen por su ubicación en la célula: ribosomas libres y ribosomas unidos a membrana. Los ribosomas libres se encuentran en el citoplasma y sirven para sintetizar proteínas solubles en agua; los ribosomas unidos a la membrana se encuentran unidos al retículo endoplásmico rugoso y producen proteínas para su inserción en la membrana celular o proteínas destinadas a la exportación de la célula.
Las diferencias entre ribosomas eucariotas y procariotas son clínicamente relevantes porque ciertos antibióticos están diseñados para dirigirse a uno u otro. Por ejemplo, la cicloheximida se dirige a la acción eucariota, mientras que el cloranfenicol se dirige a los ribosomas procariotas. 1 Dado que las células humanas son eucariotas, generalmente no se ven perjudicadas por antibióticos que destruyen los ribosomas procariotas en las bacterias. Sin embargo, a veces pueden ocurrir efectos secundarios negativos porque las mitocondrias en las células humanas contienen ribosomas procariotas.
Sistema Endomembrana
El sistema endomembrano, exclusivo de las células eucariotas, es una serie de túbulos membranosos, sacos y discos aplanados que sintetizan muchos componentes celulares y mueven materiales dentro de la célula (Figura\(\PageIndex{6}\)). Debido a su mayor tamaño celular, las células eucariotas requieren de este sistema para transportar materiales que no pueden dispersarse solo por difusión. El sistema endomembrano comprende varios orgánulos y conexiones entre ellos, incluyendo el retículo endoplásmico, aparato de Golgi, lisosomas y vesículas.
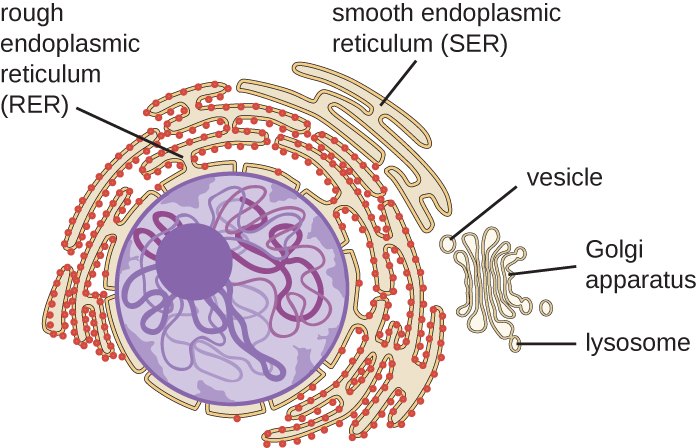
Retículo endoplásmico
El retículo endoplásmico (RE) es una matriz interconectada de túbulos y cisternas (sacos aplanados) con una sola bicapa lipídica (Figura\(\PageIndex{7}\)). Los espacios dentro de las cisternas se denominan lumen de la sala de emergencia. Hay dos tipos de ER, retículo endoplásmico rugoso (RER) y retículo endoplásmico liso (SER). Estos dos tipos diferentes de ER son sitios para la síntesis de diferentes tipos de moléculas. El RER está tachonado con ribosomas unidos en el lado citoplásmico de la membrana. Estos ribosomas producen proteínas destinadas a la membrana plasmática (Figura\(\PageIndex{}\)). Después de la síntesis, estas proteínas se insertan en la membrana del RER. Pequeños sacos del RER que contienen estas proteínas recién sintetizadas brotan como vesículas de transporte y se mueven al aparato de Golgi para su posterior procesamiento, directamente a la membrana plasmática, a la membrana de otro orgánulo, o fuera de la célula. Las vesículas transportadoras son esferas membranosas de un solo lípido, bicapa, con interiores huecos que transportan moléculas. SER no tiene ribosomas y, por lo tanto, parece “suave”. Se involucra en la biosíntesis de lípidos, metabolismo de carbohidratos y desintoxicación de compuestos tóxicos dentro de la célula.
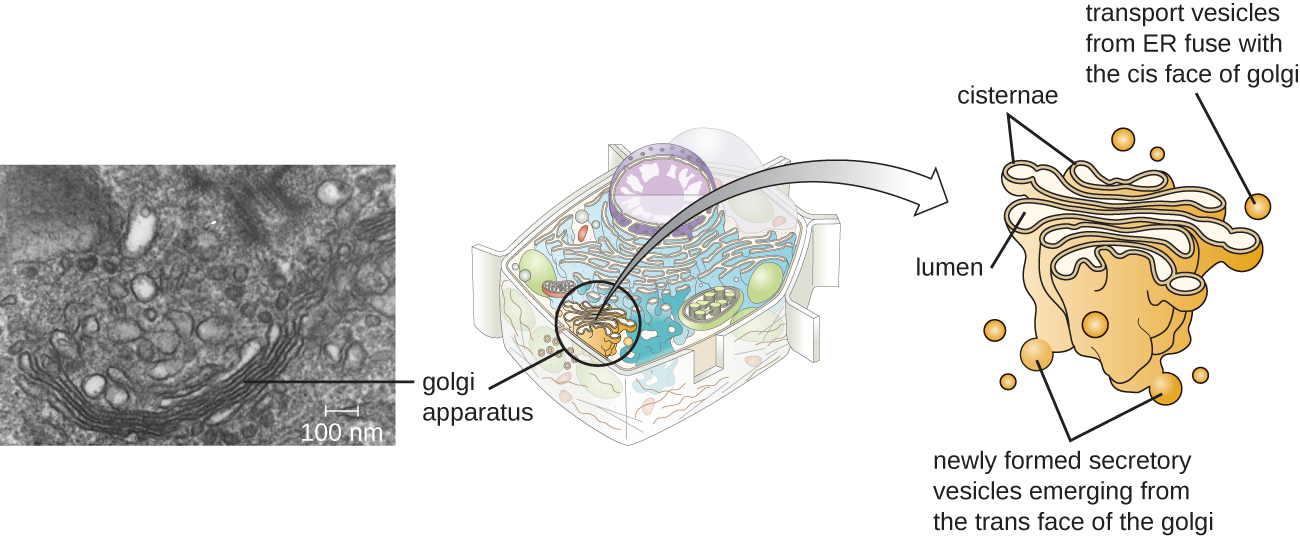
Aparato de Golgi
El aparato de Golgi fue descubierto dentro del sistema endomembrano en 1898 por el científico italiano Camillo Golgi (1843—1926), quien desarrolló una novedosa técnica de tinción que mostraba estructuras de membrana apiladas dentro de las células de Plasmodium, el agente causante de la malaria. El aparato de Golgi está compuesto por una serie de discos membranosos llamados dictyosomas, cada uno con una sola bicapa lipídica, que se apilan juntos (Figura\(\PageIndex{8}\)).
Las enzimas en el aparato de Golgi modifican los lípidos y las proteínas transportadas de la sala de emergencias al Golgi, a menudo agregándoles componentes de carbohidratos, produciendo glicolípidos, glicoproteínas o proteoglicanos. Los glicolípidos y glicoproteínas a menudo se insertan en la membrana plasmática y son importantes para el reconocimiento de señales por otras células o partículas infecciosas. Diferentes tipos de células se pueden distinguir entre sí por la estructura y disposición de los glicolípidos y glicoproteínas contenidos en sus membranas plasmáticas. Estos glicolípidos y glicoproteínas comúnmente también sirven como receptores de la superficie celular.
Las vesículas de transporte que salen del ER se fusionan con un aparato de Golgi en su cara receptora, o cis. Las proteínas se procesan dentro del aparato de Golgi, y luego vesículas de transporte adicionales que contienen las proteínas modificadas y los lípidos se pellizcan del aparato de Golgi en su cara saliente o trans. Estas vesículas salientes se mueven y se fusionan con la membrana plasmática o la membrana de otros orgánulos.
La exocitosis es el proceso por el cual las vesículas secretoras (sacos membranosos esféricos) liberan su contenido al exterior de la célula (Figura\(\PageIndex{8}\)). Todas las células tienen vías secretoras constitutivas en las que las vesículas secretoras transportan proteínas solubles que se liberan de la célula continuamente (constitutivamente). Ciertas células especializadas también tienen vías secretoras reguladas, que se utilizan para almacenar proteínas solubles en vesículas secretoras. La secreción regulada involucra sustancias que solo se liberan en respuesta a ciertos eventos o señales. Por ejemplo, ciertas células del sistema inmune humano (por ejemplo, mastocitos) secretan histamina en respuesta a la presencia de objetos extraños o patógenos en el cuerpo. La histamina es un compuesto que desencadena diversos mecanismos utilizados por el sistema inmune para eliminar patógenos.
Lisosomas
En la década de 1960, el científico belga Christian de Duve (1917-2013) descubrió lisosomas, orgánulos unidos a membrana del sistema endomembrano que contienen enzimas digestivas. Ciertos tipos de células eucariotas utilizan lisosomas para descomponer diversas partículas, como alimentos, orgánulos dañados o restos celulares, microorganismos o complejos inmunes. La compartimentación de las enzimas digestivas dentro del lisosoma permite que la célula digiera eficientemente la materia sin dañar los componentes citoplásmicos de la célula.
Ejercicio\(\PageIndex{2}\)
Nombrar los componentes del sistema endomembrano y describir la función de cada componente.
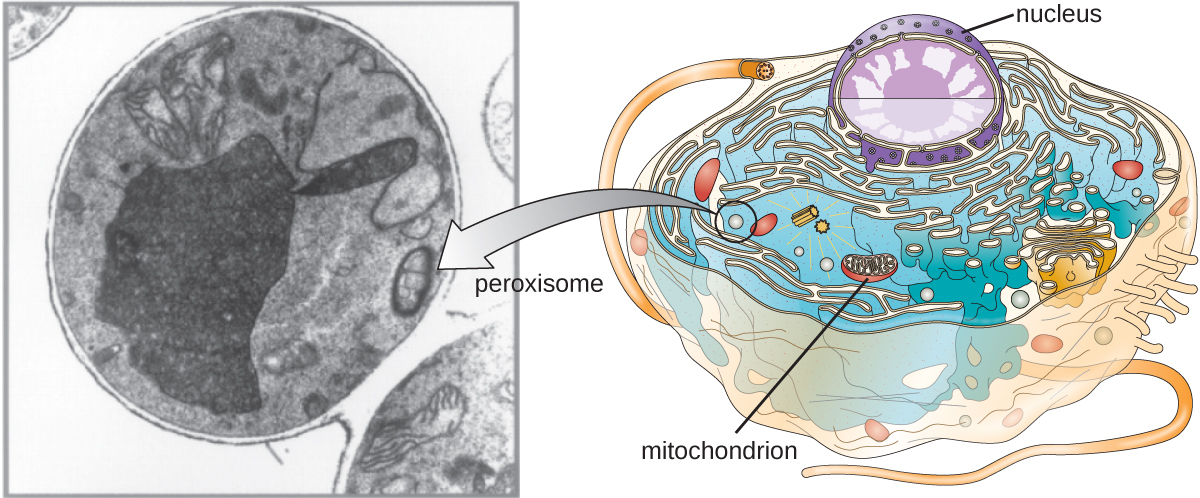
Peroxisomas
A Christian de Duve también se le atribuye el descubrimiento de peroxisomas, orgánulos unidos a membrana que no forman parte del sistema endomembrano (Figura\(\PageIndex{9}\)). Los peroxisomas se forman independientemente en el citoplasma a partir de la síntesis de proteínas peroxinas por ribosomas libres y la incorporación de estas proteínas de peroxina en peroxisomas existentes. Los peroxisomas en crecimiento luego se dividen por un proceso similar a la fisión binaria.
Los peroxisomas fueron nombrados por primera vez por su capacidad para producir peróxido de hidrógeno, una molécula altamente reactiva que ayuda a descomponer moléculas como el ácido úrico, aminoácidos y ácidos grasos. Los peroxisomas también poseen la enzima catalasa, que puede degradar el peróxido de hidrógeno. Junto con el SER, los peroxisomas también juegan un papel en la biosíntesis lipídica. Al igual que los lisosomas, la compartimentación de estas moléculas degradativas dentro de un orgánulo ayuda a proteger el contenido citoplásmico de daños no deseados.
Los peroxisomas de ciertos organismos están especializados para satisfacer sus necesidades funcionales particulares. Por ejemplo, los glioxisomas son peroxisomas modificados de levaduras y células vegetales que realizan varias funciones metabólicas, incluyendo la producción de moléculas de azúcar. De igual manera, los glucosomas son peroxisomas modificados elaborados por ciertos tripanosomas, los protozoos patógenos que causan la enfermedad de Chagas y la enfermedad africana del sueño.
Citoesqueleto
Las células eucariotas tienen un citoesqueleto interno hecho de microfilamentos, filamentos intermedios y microtúbulos. Esta matriz de fibras y tubos proporciona soporte estructural así como una red sobre la cual se pueden transportar materiales dentro de la celda y sobre la que se pueden anclar orgánulos (Figura\(\PageIndex{10}\)). Por ejemplo, el proceso de exocitosis implica el movimiento de una vesícula a través de la red citoesquelética hacia la membrana plasmática, donde puede liberar su contenido.
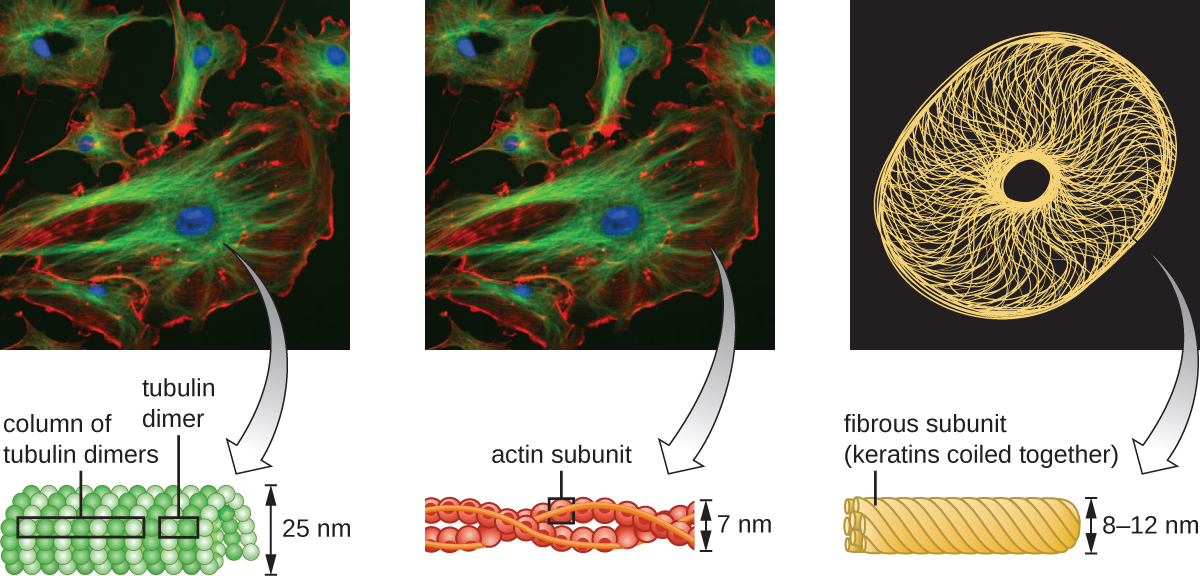
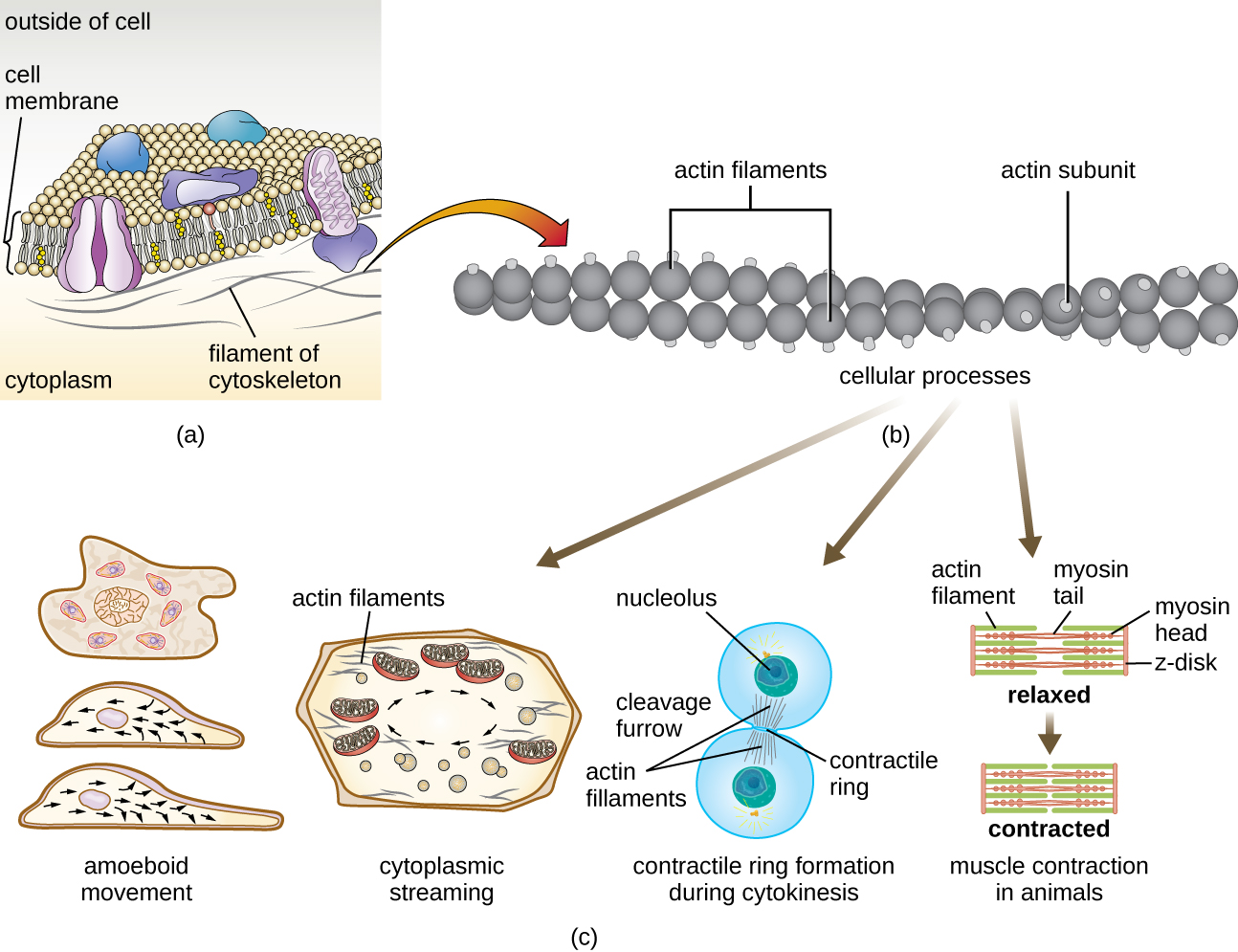
Los microfilamentos están compuestos por dos hebras entrelazadas de actina, cada una compuesta por monómeros de actina formando cables filamentosos de 6 nm de diámetro 2 (Figura\(\PageIndex{11}\)). Los filamentos de actina trabajan en conjunto con proteínas motoras, como la miosina, para efectuar la contracción muscular en animales o el movimiento ameboide de algunos microbios eucariotas. En los organismos ameboides, la actina se puede encontrar en dos formas: una forma más rígida, polimerizada, de gel y una forma más fluida, soluble no polimerizada. La actina en forma de gel crea estabilidad en el ectoplasma, el área gelatinosa del citoplasma justo dentro de la membrana plasmática de los protozoos ameboides.
Las extensiones temporales de la membrana citoplasmática llamadas pseudopodios (que significa “pies falsos”) se producen a través del flujo directo de filamentos de actina soluble hacia los pseudopodios, seguido por el ciclo gel-sol de los filamentos de actina, dando como resultado la motilidad celular. Una vez que el citoplasma se extiende hacia afuera, formando un pseudopodio, el citoplasma restante fluye hacia arriba para unirse al borde de ataque, creando así una locomoción hacia adelante. Más allá del movimiento ameboide, los microfilamentos también están involucrados en una variedad de otros procesos en las células eucariotas, incluyendo la transmisión citoplasmática (el movimiento o circulación del citoplasma dentro de la célula), la formación de surcos de escisión durante la división celular y el movimiento muscular en animales (Figura\(\PageIndex{11}\)). Estas funciones son el resultado de la naturaleza dinámica de los microfilamentos, que pueden polimerizarse y despolimerizarse con relativa facilidad en respuesta a señales celulares, y sus interacciones con motores moleculares en diferentes tipos de células eucariotas.
Los filamentos intermedios (Figura\(\PageIndex{12}\)) son un grupo diverso de filamentos citoesqueléticos que actúan como cables dentro de la célula. Se denominan “intermedios” porque su diámetro de 10 nm es más grueso que el de la actina pero más delgado que el de los microtúbulos. 3 Están compuestas por varias hebras de subunidades polimerizadas que, a su vez, están conformadas por una amplia variedad de monómeros. Los filamentos intermedios tienden a ser más permanentes en la célula y mantienen la posición del núcleo. También forman la lámina nuclear (revestimiento o capa) justo dentro de la envoltura nuclear. Adicionalmente, los filamentos intermedios juegan un papel en el anclaje de células juntas en tejidos animales. La desmina de proteína de filamento intermedio se encuentra en los desmosomas, las estructuras proteicas que unen las células musculares y las ayudan a resistir las fuerzas físicas externas. La proteína de filamento intermedio queratina es una proteína estructural que se encuentra en el cabello, la piel y las uñas.
Los microtúbulos (Figura\(\PageIndex{13}\)) son un tercer tipo de fibra citoesquelética compuesta por dímeros de tubulina (α tubulina y β tubulina). Estos forman tubos huecos de 23 nm de diámetro que se utilizan como vigas dentro del citoesqueleto. 4 Al igual que los microfilamentos, los microtúbulos son dinámicos y tienen la capacidad de ensamblar y desmontar rápidamente. Los microtúbulos también funcionan con proteínas motoras (como dineína y cinesina) para mover orgánulos y vesículas dentro del citoplasma. Adicionalmente, los microtúbulos son los componentes principales de los flagelos y cilios eucariotas, que componen tanto el filamento como los componentes basales del cuerpo (Figura\(\PageIndex{20}\)).
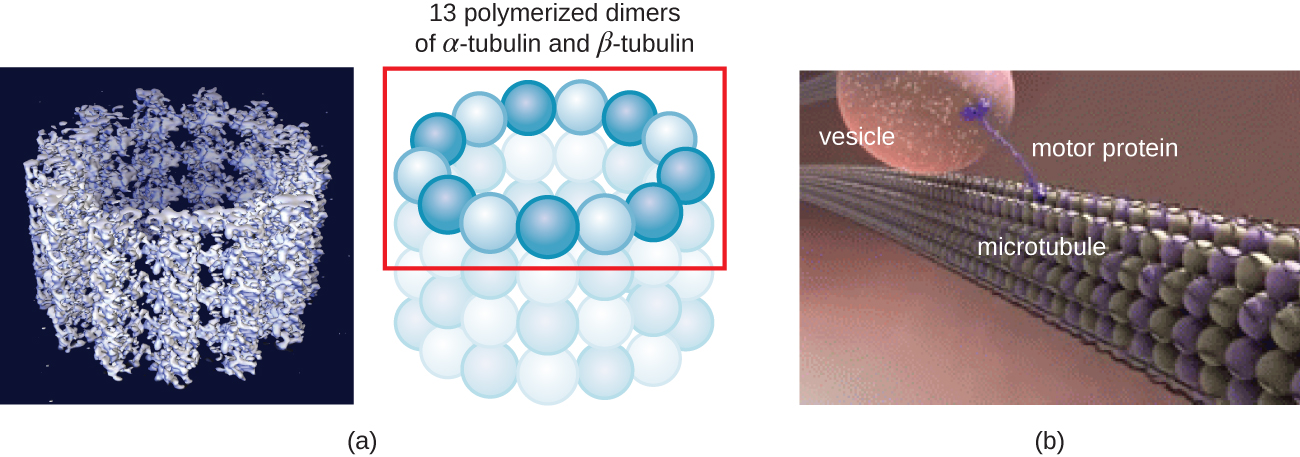
Además, los microtúbulos están involucrados en la división celular, formando el huso mitótico que sirve para separar los cromosomas durante la mitosis y meiosis. El huso mitótico es producido por dos centrosomas, que son esencialmente centros organizadores de microtúbulos, en extremos opuestos de la célula. Cada centrosoma está compuesto por un par de centriolos colocados en ángulo recto entre sí, y cada centriolo es una matriz de nueve microtúbulos paralelos dispuestos en tripletes (Figura\(\PageIndex{14}\)).
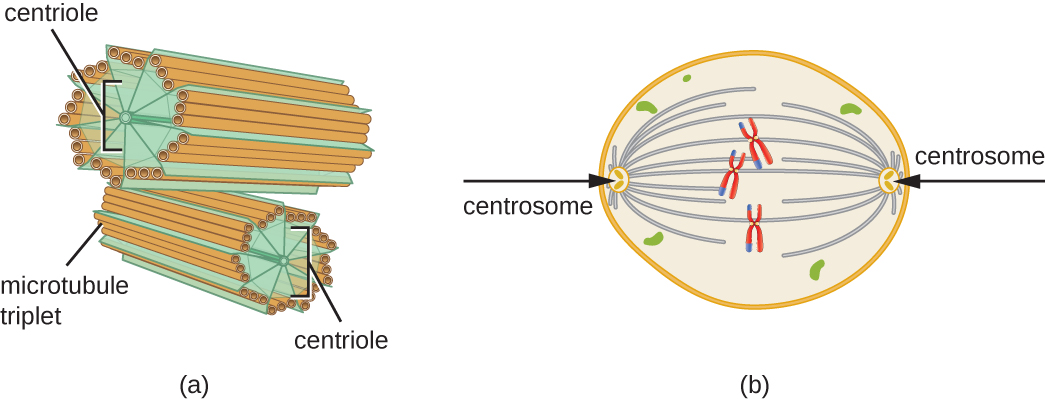
Ejercicio\(\PageIndex{3}\)
Comparar y contrastar los tres tipos de estructuras citoesqueléticas descritas en esta sección.
Mitocondrias
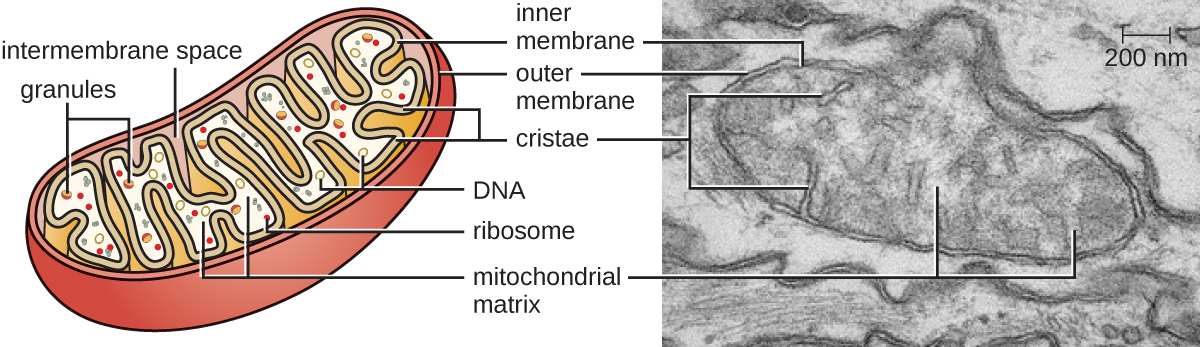
Los orgánulos grandes y complejos en los que ocurre la respiración celular aeróbica en las células eucariotas se denominan mitocondrias (Figura\(\PageIndex{15}\)). El término “mitocondria” fue acuñado por primera vez por el microbiólogo alemán Carl Benda en 1898 y posteriormente se conectó con el proceso de respiración por Otto Warburg en 1913. Los científicos durante la década de 1960 descubrieron que las mitocondrias tienen su propio genoma y ribosomas de los 70. Se encontró que el genoma mitocondrial era bacteriano, cuando se secuenció en 1976. Estos hallazgos finalmente apoyaron la teoría endosimbiótica propuesta por Lynn Margulis, que establece que las mitocondrias surgieron originalmente a través de un evento endosimbiótico en el que una bacteria capaz de respiración celular aeróbica fue absorbida por fagocitosis en una célula hospedadora y permaneció como una intracelular viable componente.
Cada mitocondria tiene dos membranas lipídicas. La membrana externa es un remanente de las estructuras de membrana de la célula hospedadora original. La membrana interna se derivó de la membrana plasmática bacteriana. La cadena de transporte de electrones para la respiración aeróbica utiliza proteínas integrales incrustadas en la membrana interna. La matriz mitocondrial, correspondiente a la ubicación del citoplasma de la bacteria original, es la ubicación actual de muchas enzimas metabólicas. También contiene ADN mitocondrial y ribosomas de los 70. Las invaginaciones de la membrana interna, llamadas cristae, evolucionaron para aumentar la superficie para la localización de reacciones bioquímicas. Los patrones de plegamiento de las cristae difieren entre varios tipos de células eucariotas y se utilizan para distinguir diferentes organismos eucariotas entre sí.
Cloroplastos
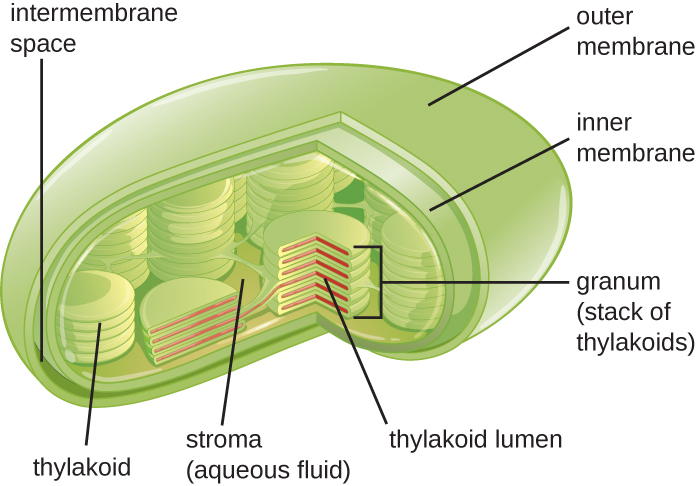
Las células vegetales y las células de algas contienen cloroplastos, los orgánulos en los que se produce la fotosíntesis (Figura\(\PageIndex{16}\)). Todos los cloroplastos tienen al menos tres sistemas de membrana: la membrana externa, la membrana interna y el sistema de membrana tilacoidea. Dentro de las membranas externa e interna se encuentra el estroma del cloroplasto, un fluido similar a un gel que constituye gran parte del volumen de un cloroplasto, y en el que flota el sistema tilacoide. El sistema tilacoide es una colección altamente dinámica de sacos de membrana plegados. Es donde se encuentra el pigmento fotosintético verde clorofila y se producen las reacciones lumínicas de la fotosíntesis. En la mayoría de los cloroplastos vegetales, los tilacoides están dispuestos en pilas llamadas grana (singular: granum), mientras que en algunos cloroplastos de algas, los tilacoides flotan libremente.
Otros orgánulos similares a las mitocondrias han surgido en otros tipos de eucariotas, pero sus papeles difieren. Los hidrogenosomas se encuentran en algunos eucariotas anaerobios y sirven como localización de la producción anaeróbica de hidrógeno. Los hidrogenosomas suelen carecer de su propio ADN y ribosomas. Los cinetoplastos son una variación de las mitocondrias que se encuentran en algunos patógenos eucariotas. En estos organismos, cada célula tiene una mitocondria única, larga y ramificada en la que el ADN de cinetoplastos, organizado como múltiples piezas circulares de ADN, se encuentra concentrado en un polo de la célula.
Organelos Relacionados con Mitocondrias en Parásitos Protozoarios
Muchos protozoos, entre ellos varios parásitos protozoarios que causan infecciones en humanos, pueden ser identificados por su apariencia inusual. Las características distintivas pueden incluir morfologías celulares complejas, la presencia de orgánulos únicos o la ausencia de orgánulos comunes. Los protozoarios Giardia lamblia y Trichomonas vaginalis son dos ejemplos.
G. lamblia, causa frecuente de diarrea en humanos y muchos otros animales, es un parásito anaerobio que posee dos núcleos y varios flagelos. Su aparato de Golgi y retículo endoplásmico están muy reducidos, y carece de mitocondrias por completo. Sin embargo, tiene orgánulos conocidos como mitosomas, orgánulos unidos a doble membrana que parecen ser mitocondrias severamente reducidas. Esto ha llevado a los científicos a creer que los antepasados de G. lamblia alguna vez poseían mitocondrias que evolucionaron hasta convertirse en mitosomas. T. vaginalis, que causa la infección de transmisión sexual vaginitis, es otro parásito protozoario que carece de mitocondrias convencionales. En cambio, posee hidrogenosomas, orgánulos ligados a la membrana doble relacionados con mitocondriales que producen hidrógeno molecular utilizado en el metabolismo celular. Los científicos creen que los hidrogenosomas, como los mitosomas, también evolucionaron a partir de las mitocondrias. 5
Membrana Plasma
La membrana plasmática de las células eucariotas es similar en estructura a la membrana plasmática procariota en que está compuesta principalmente por fosfolípidos formando una bicapa con proteínas periféricas e integrales incrustadas (Figura\(\PageIndex{17}\)). Estos componentes de la membrana se mueven dentro del plano de la membrana de acuerdo con el modelo de mosaico de fluidos. Sin embargo, a diferencia de la membrana procariota, las membranas eucariotas contienen esteroles, incluido el colesterol, que alteran la fluidez de la membrana. Además, muchas células eucariotas contienen algunos lípidos especializados, incluidos los esfingolípidos, que se cree que desempeñan un papel en el mantenimiento de la estabilidad de la membrana, además de estar involucrados en las vías de transducción de señales y la comunicación de célula a célula.

Mecanismos de transporte de membrana
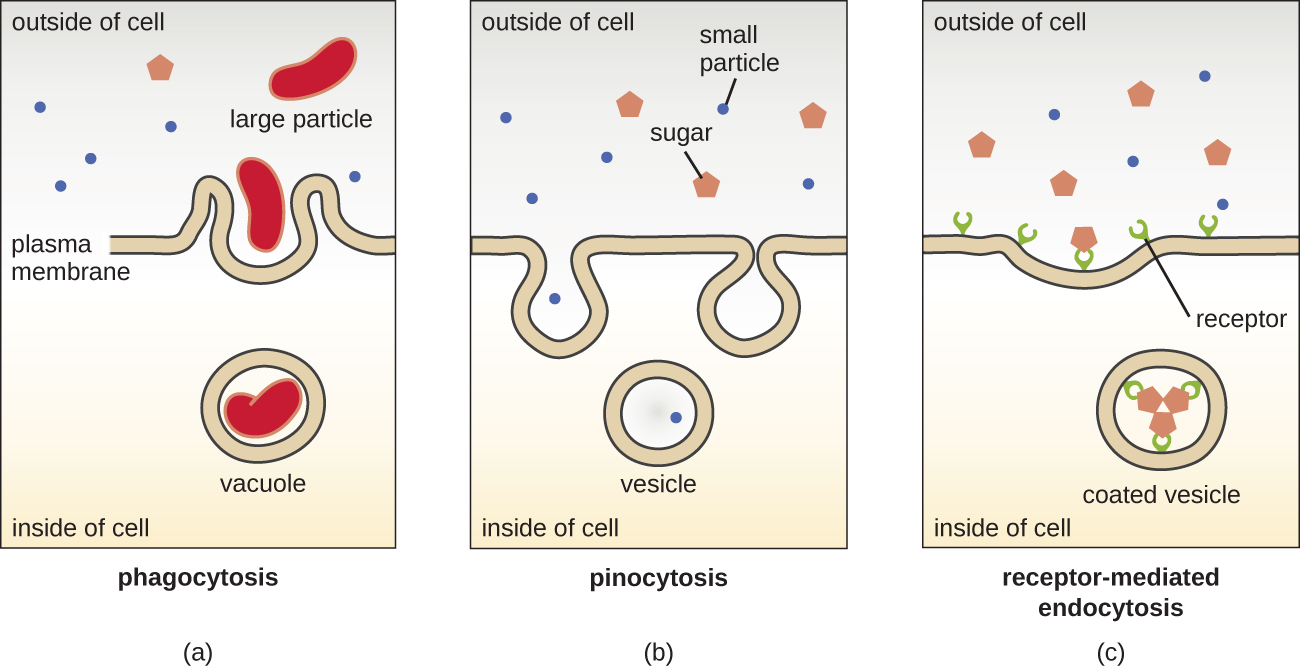
Los procesos de difusión simple, difusión facilitada y transporte activo se utilizan tanto en células eucariotas como procariotas. Sin embargo, las células eucariotas también tienen la capacidad única de realizar diversos tipos de endocitosis, la captación de materia a través de la invaginación de la membrana plasmática y la formación de vacuolas/vesículas (Figura\(\PageIndex{18}\)). Un tipo de endocitosis que implica el engullido de partículas grandes a través de la invaginación de la membrana se llama fagocitosis, que significa “alimentación celular”. En la fagocitosis, las partículas (u otras células) se encierran en un bolsillo dentro de la membrana, que luego se pellizca de la membrana para formar una vacuola que rodea completamente la partícula. Otro tipo de endocitosis se llama pinocitosis, que significa “consumo de células”. En la pinocitosis, los materiales y líquidos pequeños disueltos se llevan a la célula a través de pequeñas vesículas. Los hongos saprófitos, por ejemplo, obtienen sus nutrientes de materia muerta y en descomposición en gran parte a través de pinocitosis.
La endocitosis mediada por receptores es un tipo de endocitosis que es iniciada por moléculas específicas llamadas ligandos cuando se unen a receptores de superficie celular en la membrana. La endocitosis mediada por receptores es el mecanismo que utilizan las hormonas peptídicas y derivadas de aminas para ingresar a las células y también es utilizada por varios virus y bacterias para ingresar a las células hospedadoras.
El proceso por el cual las vesículas secretoras liberan su contenido al exterior de la célula se llama exocitosis. Las vesículas se mueven hacia la membrana plasmática y luego se funden con la membrana, expulsando su contenido fuera de la célula. La exocitosis es utilizada por las células para eliminar los productos de desecho y también puede usarse para liberar señales químicas que pueden ser absorbidas por otras células.
Pared celular
Además de una membrana plasmática, algunas células eucariotas tienen una pared celular. Las células de hongos, algas, plantas e incluso algunos protistas tienen paredes celulares. Dependiendo del tipo de célula eucariota, las paredes celulares pueden estar hechas de una amplia gama de materiales, incluyendo celulosa (hongos y plantas); sílice biogénica, carbonato de calcio, agar y carragenano (protistas y algas); o quitina (hongos). En general, todas las paredes celulares proporcionan estabilidad estructural a la célula y protección contra tensiones ambientales como la desecación, cambios en la presión osmótica y lesiones traumáticas. 6
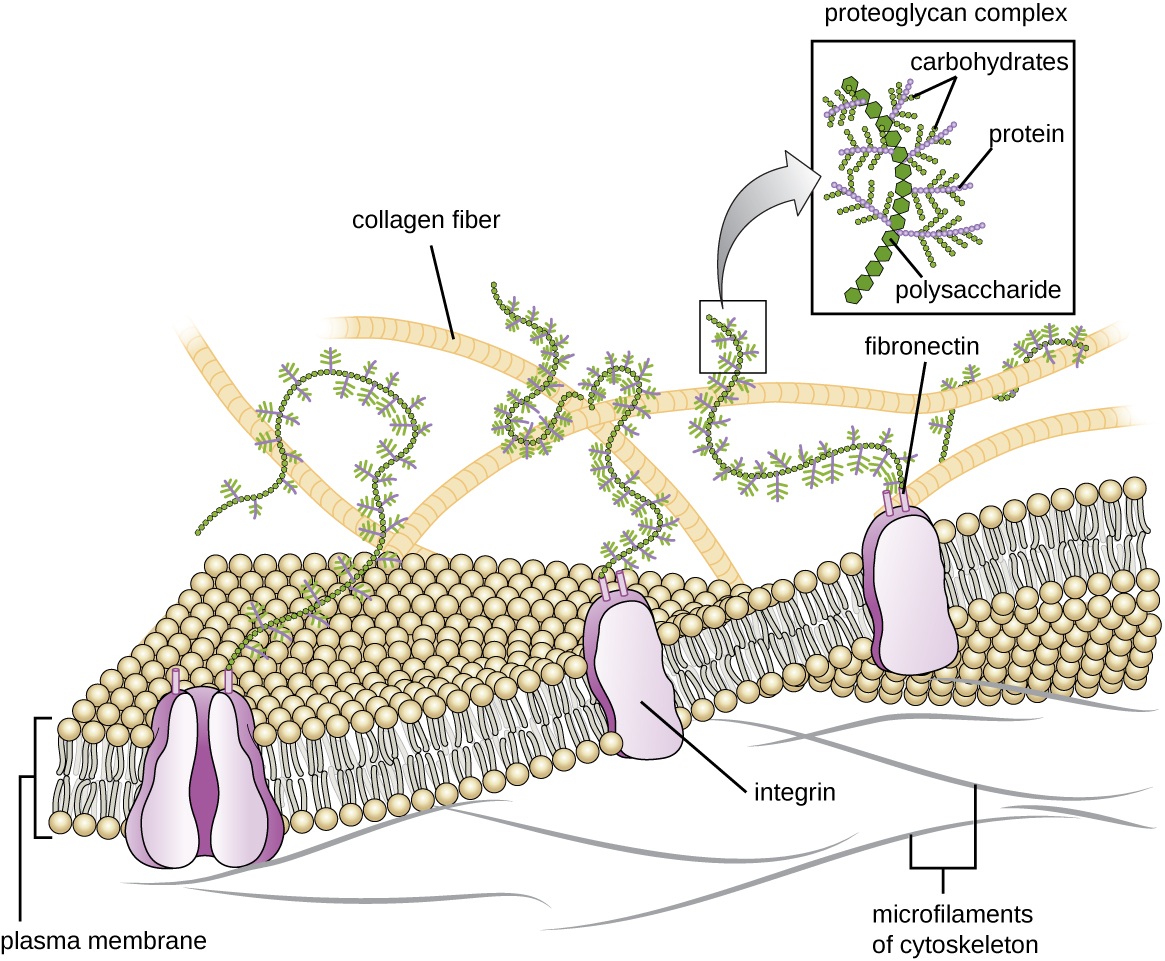
Matriz extracelular
Las células de animales y algunos protozoos no tienen paredes celulares para ayudar a mantener la forma y proporcionar estabilidad estructural. En cambio, estos tipos de células eucariotas producen una matriz extracelular para este propósito. Secretan una masa pegajosa de carbohidratos y proteínas en los espacios entre las células adyacentes (Figura\(\PageIndex{19}\)). Algunos componentes proteicos se ensamblan en una membrana basal a la que se adhieren los componentes restantes de la matriz extracelular. Los proteoglicanos típicamente forman la masa voluminosa de la matriz extracelular mientras que las proteínas fibrosas, como el colágeno, proporcionan fuerza. Tanto los proteoglicanos como el colágeno están unidos a las proteínas de fibronectina, las cuales, a su vez, se unen a las proteínas integrinas. Estas proteínas integrinas interactúan con proteínas transmembrana en las membranas plasmáticas de células eucariotas que carecen de paredes celulares.
En las células animales, la matriz extracelular permite que las células dentro de los tejidos soporten tensiones externas y transmitan señales desde el exterior de la célula hacia el interior. La cantidad de matriz extracelular es bastante extensa en varios tipos de tejidos conectivos, y las variaciones en la matriz extracelular pueden dar a diferentes tipos de tejidos sus distintas propiedades. Además, la matriz extracelular de una célula hospedadora suele ser el sitio donde los patógenos microbianos se unen para establecer la infección. Por ejemplo, Streptococcus pyogenes, la bacteria que causa la faringitis estreptocócica y varias otras infecciones, se une a la fibronectina en la matriz extracelular de las células que recubren la orofaringe (región superior de la garganta).
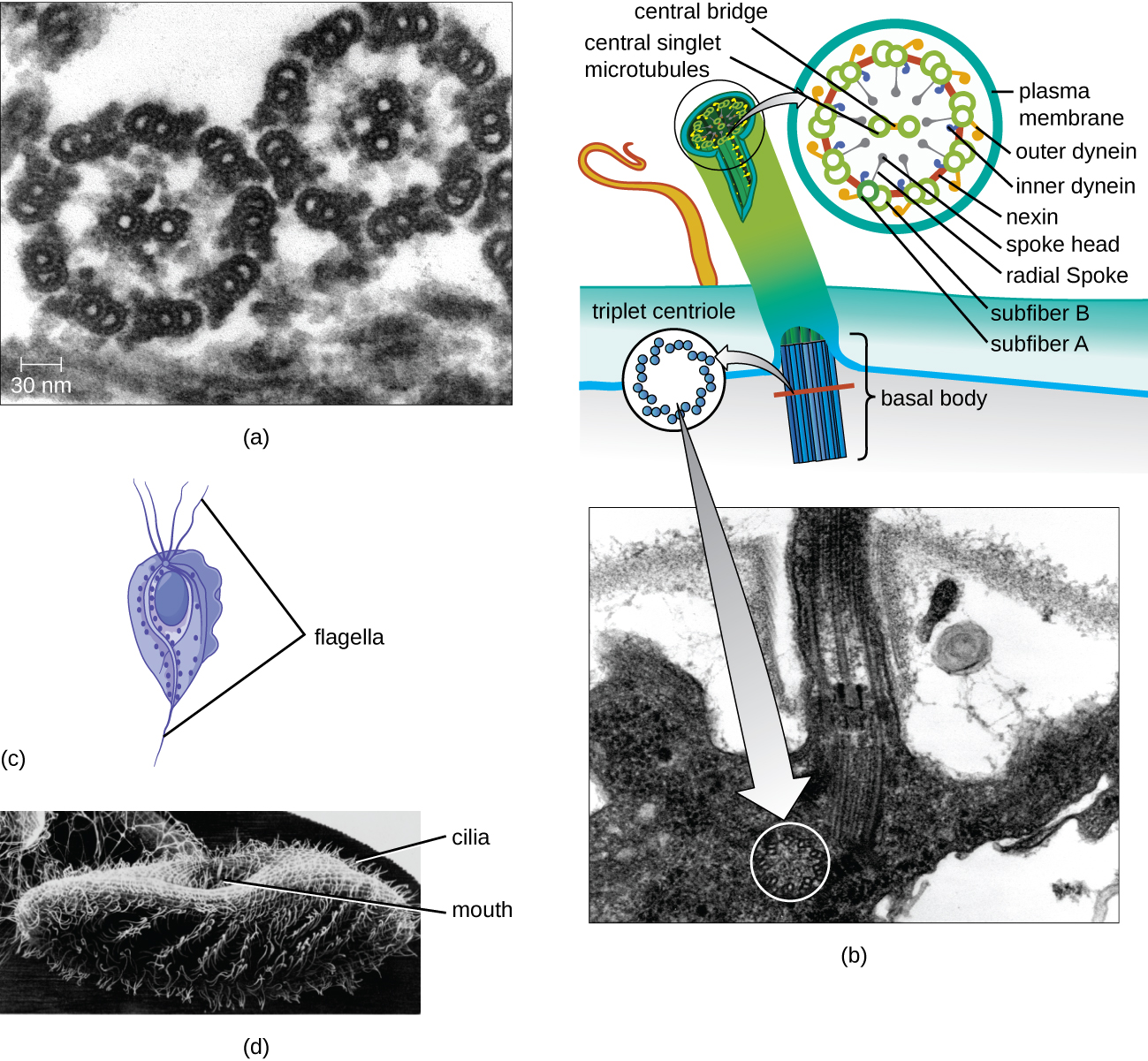
Flagelos y Cilios
Algunas células eucariotas utilizan flagelos para la locomoción; sin embargo, los flagelos eucariotas son estructuralmente distintos de los que se encuentran en las células procariotas. Mientras que el flagelo procariota es una estructura rígida y giratoria, un flagelo eucariota es más como un látigo flexible compuesto por nueve pares paralelos de microtúbulos que rodean un par central de microtúbulos. Esta disposición se conoce como una matriz 9+2 (Figura\(\PageIndex{20}\)). Los microtúbulos paralelos utilizan proteínas motoras dineínas para moverse entre sí, haciendo que el flagelo se doble.
Los cilios (singular: cilio) son una estructura externa similar que se encuentra en algunas células eucariotas. Exclusivos de los eucariotas, los cilios son más cortos que los flagelos y a menudo cubren toda la superficie de una célula; sin embargo, son estructuralmente similares a los flagelos (una matriz 9+2 de microtúbulos) y utilizan el mismo mecanismo para el movimiento. Una estructura llamada cuerpo basal se encuentra en la base de cada cilio y flagelo. El cuerpo basal, que une el cilio o flagelo a la célula, está compuesto por una matriz de microtúbulos tripletes similares a la de un centriolo pero incrustados en la membrana plasmática. Debido a su longitud más corta, los cilios utilizan un movimiento rápido, flexible y ondulante. Además de la motilidad, los cilios pueden tener otras funciones como barrer partículas más allá o hacia las células. Por ejemplo, los protozoos ciliados utilizan el barrido de los cilios para mover partículas de alimentos a sus partes bucales, y las células ciliadas en el tracto respiratorio de los mamíferos laten en sincronía para barrer el moco y los desechos hacia arriba y fuera de los pulmones (Figura\(\PageIndex{20}\)).
Ejercicio\(\PageIndex{4}\)
- Explicar cómo se compara la envoltura celular de las células eucariotas con la de las células procariotas.
- Explicar la diferencia entre flagelos eucariotas y procariotas.
Foco Clínico: Resolución
Dado que la amoxicilina no ha resuelto el caso de neumonía de Barbara, la AP prescribe otro antibiótico, la azitromicina, que se dirige a los ribosomas bacterianos en lugar de peptidoglicano. Después de tomar la azitromicina como se le indica, los síntomas de Barbara se resuelven y finalmente comienza a sentirse como ella misma otra vez. Al presumir que no hubo resistencia farmacológica a la amoxicilina, y dada la efectividad de la azitromicina, el agente causante de la neumonía de Bárbara es probablemente Mycoplasma pneumoniae. A pesar de que esta bacteria es una célula procariota, no es inhibida por la amoxicilina porque no tiene una pared celular y, por lo tanto, no produce peptidoglicano.
Conceptos clave y resumen
- Las células eucariotas se definen por la presencia de un núcleo que contiene el genoma de ADN y unido por una membrana nuclear (o envoltura nuclear) compuesta por dos bicapas lipídicas que regulan el transporte de materiales dentro y fuera del núcleo a través de los poros nucleares.
- Las morfologías de las células eucariotas varían mucho y pueden mantenerse mediante diversas estructuras, incluyendo el citoesqueleto, la membrana celular y/o la pared celular.
- El nucleolo, ubicado en el núcleo de las células eucariotas, es el sitio de síntesis ribosómica y las primeras etapas del ensamblaje de ribosomas.
- Las células eucariotas contienen ribosomas 80S en el retículo endoplásmico rugoso (ribosomas unidos a la membrana) y citoplasma (ribosomas libres). Contienen ribosomas de los años 70 en mitocondrias y cloroplastos.
- Las células eucariotas han desarrollado un sistema endomembrano, que contiene orgánulos unidos a membrana involucrados en el transporte. Estos incluyen vesículas, el retículo endoplásmico y el aparato de Golgi.
- El retículo endoplásmico liso juega un papel en la biosíntesis lipídica, el metabolismo de carbohidratos y la desintoxicación de compuestos tóxicos. El retículo endoplásmico rugoso contiene ribosomas 80S unidos a membrana que sintetizan proteínas destinadas a la membrana celular
- El aparato de Golgi procesa proteínas y lípidos, típicamente a través de la adición de moléculas de azúcar, produciendo glicoproteínas o glicolípidos, componentes de la membrana plasmática que se utilizan en la comunicación célula a célula.
- Los lisosomas contienen enzimas digestivas que descomponen pequeñas partículas ingeridas por endocitosis, partículas grandes o células ingeridas por fagocitosis y componentes intracelulares dañados.
- El citoesqueleto, compuesto por microfilamentos, filamentos intermedios y microtúbulos, proporciona soporte estructural en células eucariotas y sirve como red para el transporte de materiales intracelulares.
- Los centrosomas son centros organizadores de microtúbulos importantes en la formación del huso mitótico en la mitosis.
- Las mitocondrias son el sitio de respiración celular. Tienen dos membranas: una membrana externa y una interna con cristae. La matriz mitocondrial, dentro de la membrana interna, contiene el ADN mitocondrial, los ribosomas de los 70 y las enzimas metabólicas.
- La membrana plasmática de las células eucariotas es estructuralmente similar a la que se encuentra en las células procariotas, y los componentes de la membrana se mueven según el modelo de mosaico de fluidos. Sin embargo, las membranas eucariotas contienen esteroles, que alteran la fluidez de la membrana, así como glicoproteínas y glicolípidos, que ayudan a la célula a reconocer otras células y partículas infecciosas.
- Además del transporte activo y el transporte pasivo, las membranas celulares eucariotas pueden llevar material a la célula a través de endocitosis, o expulsar materia de la célula a través de exocitosis.
- Las células de hongos, algas, plantas y algunos protistas tienen una pared celular, mientras que las células de animales y algunos protozoos tienen una matriz extracelular pegajosa que proporciona soporte estructural y media la señalización celular.
- Los flagelos eucariotas son estructuralmente distintos de los flagelos procariotas pero tienen un propósito similar (locomoción). Los cilios son estructuralmente similares a los flagelos eucariotas, pero más cortos; pueden usarse para la locomoción, alimentación o movimiento de partículas extracelulares.
Notas al pie
- 1 A.E. Barnhill, M.T. Brewer, S.A. Carlson. “Efectos adversos de los antimicrobianos a través de la inhibición predecible o idiosincrásica de los componentes mitocondriales del huésped”. Agentes Antimicrobianos y Quimioterapia 56 núm. 8 (2012) :4046—4051.
- 2 Fuchs E, Cleveland DW. “Un Andamiaje Estructural de Filamentos Intermedios en Salud y Enfermedad”. Ciencia 279 núm. 5350 (1998) :514—519.
- 3 E. Fuchs, D.W. Cleveland. “Un Andamiaje Estructural de Filamentos Intermedios en Salud y Enfermedad”. Ciencia 279 núm. 5350 (1998) :514—519.
- 4 E. Fuchs, D.W. Cleveland. “Un Andamiaje Estructural de Filamentos Intermedios en Salud y Enfermedad”. Ciencia 279 núm. 5350 (1998) :514—519.
- 5 N. Yarlett, J.H.P. Hackstein. “Hidrogenosomas: Un Organelo, Múltiples Orígenes”. BioScience 55 núm. 8 (2005) :657—658.
- 6 M. Dudzick. “Protistas”. OpenStax CNX. 27 de noviembre de 2013. http://cnx.org/contents/f7048bb6-e46...ef291cf7049c@1