
10.4: Mecanismos sinápticos de aprendizaje y memoria
- Page ID
- 148185
Objetivos de aprendizaje
- Describir la teoría del engrama de Hebb
- Definir la regla de Hebb
- Definir sinapsis Hebb y conjuntos de celdas
- Describir los hallazgos de Kandel sobre los cambios sinápticos que median la habituación y sensibilización en la babosa marina, Aplysia
- Explicar la teoría de doble traza de la memoria
- Describir el papel del hipocampo en el aprendizaje y la memoria
- Describir el papel del cerebelo y la amígdala en el aprendizaje y la memoria
- Discutir Potenciación a Largo Plazo (LTP) y Depresión a largo plazo y aprendizaje y memoria
- Describir los cambios en las espinas dendríticas asociados con el aprendizaje y la memoria
- Discutir cómo la LTP y los cambios anatómicos en las espinas sinápticas podrían estar relacionados con la teoría de doble traza de Hebb
Visión general
La teoría de doble traza de Donald Hebb (1949) de la base física de la memoria se centró en los cambios en la sinapsis como base para el aprendizaje y la memoria. Esta teoría influyente condujo al descubrimiento de cambios en la liberación de transmisores asociados con el aprendizaje en el sistema nervioso invertebrado de la babosa marina. Otros investigadores encontraron aumentos sostenidos en la conductancia sináptica causados por la estimulación de alta frecuencia de la neurona presináptica (Potenciación a Largo Plazo, LTP) así como una disminución sostenida de la fuerza sináptica conocida como Depresión a Largo Plazo (LTD). Los investigadores también han descubierto cambios anatómicos en las espinas dendríticas asociados con LTP y aprendizaje y memoria, sugiriendo cambios anatómicos en las sinapsis como posible mecanismo de aprendizaje y memoria a largo plazo a nivel sináptico. Si bien los primeros estudios en busca del “engrama”, la base física para el aprendizaje y la memoria, concluyeron que estas funciones estaban ampliamente distribuidas en todo el cerebro, investigaciones posteriores encontraron que el hipocampo es crítico para la formación de recuerdos explícitos a largo plazo y que el cerebelo está involucrado en memoria implícita, cuando la amígdala juega un papel esencial en los recuerdos emocionales.
La búsqueda del aprendizaje y la memoria en la sinapsis
por Kenneth A. Koenigshofer, Ph.D.
Así como la información se almacena en medios digitales como DVD, discos duros y unidades flash, la información en nuestra memoria a largo plazo debe almacenarse físicamente en el cerebro. Según la teoría actual, la capacidad de mantener la información en la memoria a largo plazo implica un fortalecimiento gradual de las conexiones sinápticas entre las neuronas. Cuando las vías en estas redes neuronales se activan frecuente y repetidamente, las sinapsis se vuelven más eficientes permitiendo una comunicación mejorada entre las neuronas de la red, y estos cambios crean memoria (Fundación Saylor, 2012). Como veremos, esta visión de la base física de la memoria estuvo fuertemente influenciada por las ideas de un psicólogo biológico de Harvard llamado Donald Hebb.
Hace más de un siglo un fisiólogo ruso llamado Ivan Pavlov propuso una teoría de cómo se producía el aprendizaje en el cerebro. Como recuerdas del módulo 10.2, Pavlov descubrió el condicionamiento clásico cuando observó que los repetidos emparejamientos de una campana y comida finalmente llevaron a los perros a salivar al sonido de la campana, incluso cuando la comida no estaba presente. Para explicar este fenómeno, Pavlov planteó la hipótesis de que el estímulo condicionado (CS), una campana en este caso, generó un locus de actividad neuronal en la corteza sensorial (corteza auditiva en este experimento) que irradiaba hacia afuera sobre la superficie cortical. Esto fue seguido por un locus similar de actividad neuronal cortical generado por el estímulo no condicionado (US) (carne en polvo en los experimentos clásicos de Pavlov) que también configuró ondas de actividad neural radiante. Pavlov propuso que estos dos campos en expansión de actividad neuronal, originados en diferentes áreas de la corteza, se cruzarían entre sí. Según Pavlov, esta intersección de los campos corticales de actividad neural, generados por el CS y US, formó la base neurológica para la asociación entre CS y US.
Esta teoría temprana de las bases físicas del aprendizaje y la memoria fue puesta a prueba por el psicólogo experimental, Karl Lashley, de Harvard y posteriormente, el Laboratorio Yerkes de Biología de Primates. Lashley realizó cortes entrecruzados a lo largo de la corteza cerebral de ratas y luego intentó acondicionarlas. Lo hizo para interrumpir cualquier actividad neuronal radiante en la corteza que pudiera estar presente durante el acondicionamiento, como lo había propuesto Pavlov. Lashley encontró que las ratas aún podían ser acondicionadas y que retuvieron la respuesta condicionada más tarde a pesar de los cortes entrecruzados en toda su corteza cerebral. Esto desmintió la teoría de Pavlov de que la base física del aprendizaje de una asociación por condicionamiento clásico era la intersección de ondas de actividad neural que irradiaban hacia afuera desde loci corticales.
Lashley, al igual que Pavlov, estaba interesado en encontrar el “engrama”. Este término se refiere a la traza de memoria física, la representación neuronal de la memoria en el cerebro. Hacia este fin, Lashley realizó experimentos adicionales en los que entrenó a ratas para navegar por un laberinto y luego destruyó diferentes partes de sus cerebros. Encontró que sin importar dónde se ubicara el daño cerebral, las ratas aún conservaban algún recuerdo del laberinto (Lashley, 1929, 1943, 1950). Lashley interpretó estos resultados como evidencia de que fue la cantidad de tejido cortical extirpado, no su ubicación, lo que determinó el grado de deterioro en el aprendizaje y la memoria. También planteó la hipótesis de que los recuerdos estaban ampliamente distribuidos por todo el cerebro y que por lo tanto no había un área particular del cerebro que fuera especialmente crítica para la formación de la memoria.
Lashley propuso dos principios derivados de su investigación sobre la base física del aprendizaje y la memoria: 1) acción masiva (la cantidad de corteza destruida determina el grado de deterioro en el aprendizaje y la memoria) y 2) equipotencialidad (cualquier parte de la corteza dentro de un área funcional puede hacerse cargo de las funciones de cualquier otra parte de esa misma área funcional).
Las conclusiones de Lashley tuvieron dos impactos primarios. Primero, su análisis “tendió a desacreditar, o al menos desenfatizar, el papel de las partes interiores [subcorticales] del cerebro en el aprendizaje y la memoria y, por otro, tendió a localizar los mecanismos de aprendizaje y memoria dentro de los confines de la corteza cerebral” (Thompson, 1974). En segundo lugar, los principios de acción de masas y equipotencialidad de Lashley llevaron a un sesgo anti-localización en el estudio del aprendizaje y la memoria, lo que desalentó los intentos de encontrar estructuras cerebrales específicas involucradas en estas funciones. Sin embargo, de acuerdo con los puntos de vista actuales, y contradiciendo los hallazgos de Lashley, Kaada, Rasmussen y Kveim (1961) informaron que las lesiones del sistema hipocampo-fórnix alteraron el rendimiento del laberinto. Este fue un estudio temprano que confirma lo que conocemos hoy, que una estructura subcortical, el hipocampo, es fundamental para muchas formas de aprendizaje y memoria.
Regla de Hebb, conjuntos de celdas y el engrama
Después de recibir su doctorado de Harvard en 1936, uno de los estudiantes de posgrado de Lashley, Donald Hebb, publicó T h e Organization of Behavior: A Neuropsychological Theory. En este influyente libro, Hebb propuso que el “engrama” consistió en cambios en las sinapsis durante el aprendizaje y la formación de la memoria. Una cita del libro (Hebb, 1949, p. 62) explica el principio central de Hebb del cambio sináptico en el aprendizaje y la memoria:
“Cuando un axón de la célula A está lo suficientemente cerca como para excitar una célula B y de manera repetida o persistente participa en su disparo, se produce algún proceso de crecimiento o cambio metabólico en una o ambas células de tal manera que aumenta la eficiencia de A, como una de las células que disparan B”.
Aquí Hebb está proponiendo un aumento en la fuerza de las conexiones sinápticas entre neuronas que constantemente se disparan juntas y que esto implica crecimiento o eventos metabólicos en una o ambas células.
Más simplemente declaró: “Neuronas que disparan juntas, se conectan juntas”. Esto se conoce como "regla de Hebb" (recuperado de https://can-acn.org/donald-olding-hebb/ 8 de agosto de 2021).
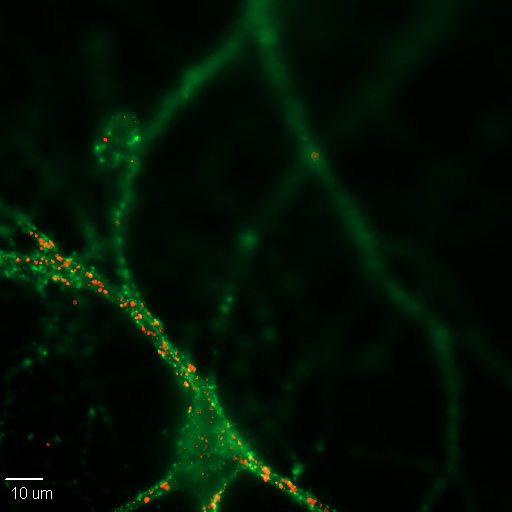
Figura\(\PageIndex{1}\): Imagen 20X de una neurona cortical de ratón cultivada en cultivo celular. Las sinapsis están marcadas para proteínas pre- (verdes) y possinápticas (rojas), sinaptofisina y PSD-95, respectivamente. (Imagen y subtítulo de Wikimedia, Synapse; https://commons.wikimedia.org/wiki/F...n_Synapses.jpg; autor, Dchordpdx; licenciado bajo la licencia Creative Commons Attribution 4.0 International).
Los neurocientíficos utilizan el término, sinapsis hebbianas, para referirse a las sinapsis que siguen este principio. Hebb también propuso la idea de “conjuntos de celdas”. Hebb planteó la hipótesis de que cuando las células se disparan juntas repetidamente, cableadas juntas, eventualmente formarían estructuras más grandes, “ensamblajes celulares”, que formarían representaciones neuronales de percepciones completas y complejas, ideas, recuerdos y otras estructuras cognitivas, como esquemas, y altamente abstractas categorías y conceptos. Todo esto podría construirse a partir de cambios en la eficiencia de la transmisión sináptica en sinapsis específicas, siguiendo el principio de Hebb de que “las neuronas que disparan juntas, se conectan”. Uno puede imaginar que conjuntos celulares enteros que se disparan juntos también podrían conectarse, creando representaciones neuronales mucho más grandes y mucho más complejas en el cerebro de ideas complejas y abstractas que incluyen cosas como teorías científicas y formulaciones matemáticas.
Aprendizaje No Asociado en Aplysia
Estas ideas han inspirado la búsqueda por parte de los neurocientíficos de cambios en las sinapsis y en las neuronas pre o possinápticas, durante y después del aprendizaje, cambios que podrían ser la base física en el sistema nervioso para el aprendizaje y la memoria (el “engrama” de Lashley, que, por cierto, no tiene nada que ver con lo no científico uso del término por parte de Scientology). Gran parte del trabajo científico pionero en la búsqueda del “engrama” ha implicado la búsqueda de estos cambios físicos asociados con el aprendizaje en el sistema nervioso simple de una especie marina, Aplysia californica, un tipo de babosa marina. Una razón importante para la selección de esta especie para su estudio, aparte de la simplicidad de su sistema nervioso (solo 20,000 neuronas), es el gran tamaño de sus neuronas lo que facilita el trabajo y las observaciones.

Figura\(\PageIndex{2}\): Aplysia californica. (Imagen de Wikimedia, Aplysia californica; https://commons.wikimedia.org/wiki/F... , _Monterey.jpg; Chad King/NOAA MBNMS; esta imagen es de dominio público porque contiene materiales que originalmente provinieron de la Administración Nacional Oceánica y Atmosférica de Estados Unidos, tomados o hechos como parte del funcionario de un empleado deberes).
Eric Kandel (1976) y sus colegas realizaron un trabajo pionero que mostró cambios sinápticos que median la habituación (ver secciones anteriores) del sifón de Aplysia y el reflejo de retirada defensiva branquial. Se encontró una disminución de la conductividad en las sinapsis involucradas en el sifón y reflejo de abstinencia branquial en Aplysia después de la habituación a una presentación repetida de un nuevo estímulo inofensivo. Kandel y sus colaboradores mostraron que la neurona sensorial presináptica liberó menos neurotransmisor sobre la neurona motora postsináptica como resultado de la presentación repetida del nuevo estímulo que condujo a la habituación del reflejo. Se encontró lo contrario para una mayor capacidad de respuesta. Sensibilización se refiere a la mejora de la capacidad de respuesta a un estímulo familiar. Una sola descarga eléctrica pequeña en la cola de Aplysia aumenta su respuesta de retirada branquial durante minutos a horas. Kandel y sus colegas encontraron que la sensibilización estaba mediada por un aumento en la liberación de neurotransmisor de la neurona sensorial presináptica a la neurona motora possináptica que sirve al músculo branquial. Así, la sensibilización es lo opuesto a la habituación tanto conductual como a nivel de liberación de transmisores (Kolb y Whishaw, 2001). Estudios posteriores mostraron cambios sinápticos que implicaban la liberación modificada del transmisor durante el acondicionamiento clásico también (Kandel y Schwartz, 1982).
Teoría de la memoria de doble rastro de Hebb
Hebb también propuso una teoría de doble traza de la memoria, un proceso eléctrico dinámico a corto plazo para la memoria a corto plazo (Hebb se refirió a “circuitos de reverberación” que sostienen esta breve actividad eléctrica) y, después de algún tiempo después, un cambio estructural duradero en sinapsis como base física (el engrama) para la memoria a largo plazo (Hebb, 1949). Siguieron décadas de investigación utilizando diversos agentes, entre ellos el choque electroconvulsivo (ECS), poco después de aprender en ratas a interrumpir el rastro de memoria a corto plazo, que según Hebb, consistía en circular o “reverberar” la actividad eléctrica, impidiendo así el almacenamiento a largo plazo. Sin embargo, aunque el ECS en ratas dentro de aproximadamente 30 segundos de aprendizaje pareció interrumpir la consolidación del rastro de memoria a corto plazo en un rastro permanente a largo plazo como lo demuestra la aparente amnesia para la tarea de aprendizaje, investigaciones adicionales sugirieron explicaciones alternativas para estos resultados. Una alternativa fue que el ECS no impidió la consolidación de la traza de memoria a largo plazo, sino que interrumpió la recuperación de la memoria. La evidencia de esta hipótesis de falla de recuperación sugiere que el fracaso original de la interpretación de consolidación de la investigación ECS probablemente fue incorrecto (Miller y Martin, 2014).
Los papeles especiales del hipocampo, el cerebelo y la amígdala
Como se discutió anteriormente, la investigación desde Lashley ha revelado que su especulación de que todas las áreas del cerebro estaban igualmente involucradas en el aprendizaje y la memoria era incorrecta. Ahora sabemos que una de las regiones cerebrales más importantes en la memoria explícita es el hipocampo, que sirve como preprocesador y elaborador de información (Squire, 1992). El hipocampo nos ayuda a codificar información sobre las relaciones espaciales, el contexto en el que se vivieron los eventos y las asociaciones entre memorias (Eichenbaum, 1999). El hipocampo también sirve en parte como un punto de conmutación que retiene la memoria por un corto tiempo y luego dirige la información a otras partes del cerebro, como la corteza, para hacer realmente el ensayo, elaboración y almacenamiento a largo plazo (Jonides, Lacey, & Nee, 2005; Fundación Saylor, 2015). También ahora sabemos que diferentes partes del cerebro están involucradas en diferentes tipos de memoria.
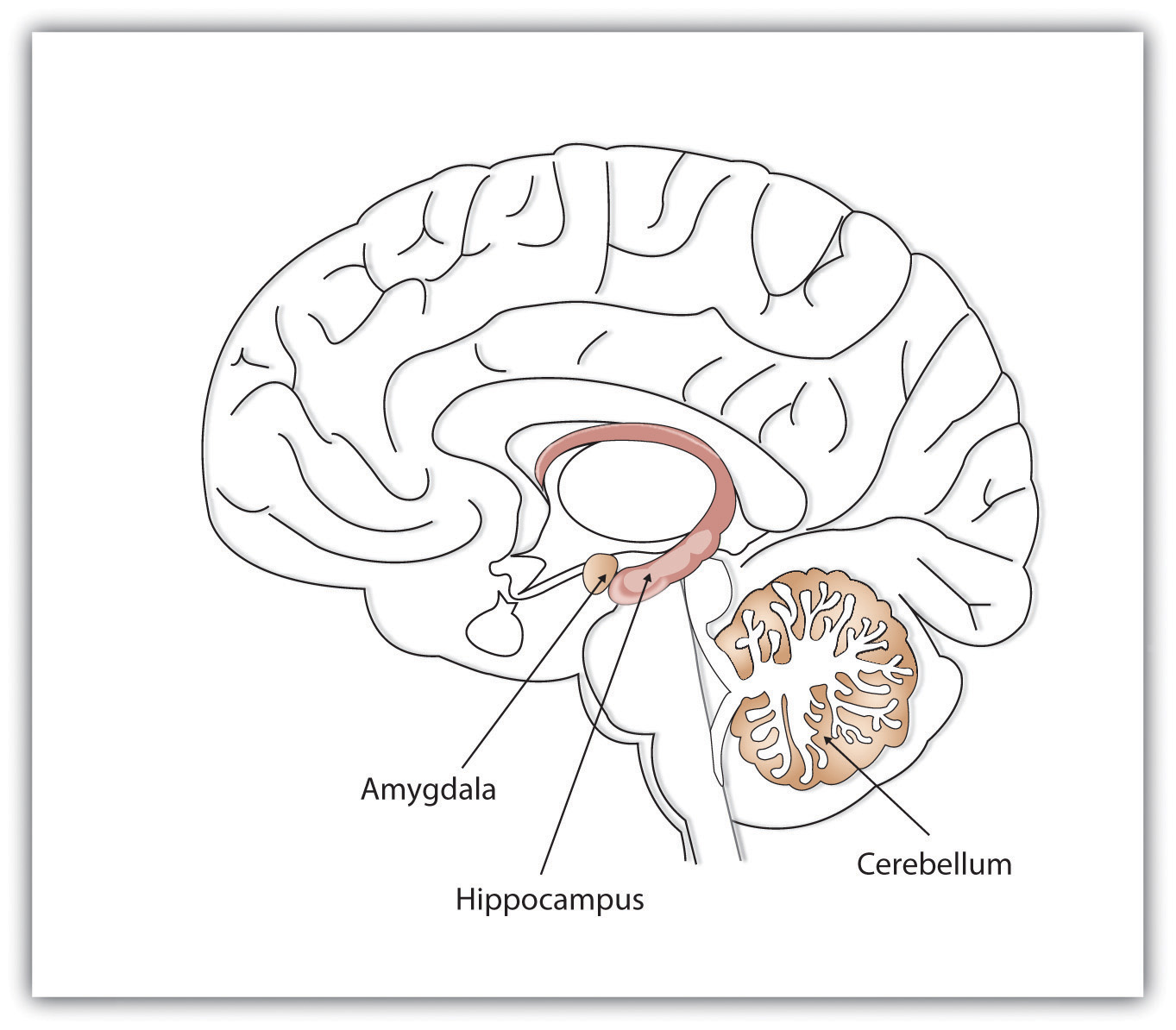
Figura\(\PageIndex{3}\): Diferentes estructuras cerebrales nos ayudan a recordar diferentes tipos de información. El hipocampo es particularmente importante en las memorias explícitas, el cerebelo en las memorias implícitas y la amígdala en las memorias emocionales (Imagen y pie de foto de la Fundación Saylor, 2015; Recordando y Juzgando; https://learn.umgc.edu/d2l/le/conten.../25917628/View; licenciado bajo una Licencia Creative Commons Attribution 3.0 Unported.).
Mientras que el hipocampo está manejando la memoria explícita, el cerebelo y la amígdala están involucrados críticamente en recuerdos implícitos y emocionales, respectivamente. La investigación muestra que el cerebelo es más activo cuando estamos aprendiendo asociaciones y en tareas de cebado (basadas en la memoria implícita), y los animales y humanos con daño al cerebelo se ven afectados en el condicionamiento clásico (Krupa, Thompson, & Thompson, 1993; Woodruff-Pak, Goldenberg, Downey-Lamb, Boyko, & amp; Lemieux, 2000). El almacenamiento de muchos de nuestros recuerdos emocionales más importantes, y particularmente los relacionados con el miedo, es iniciado y controlado por la amígdala (Sigurdsson, Doyère, Cain, & LeDoux, 2007).
Cambios en la sinapsis correlacionados con el aprendizaje y la memoria
La teoría de Hebb de que la memoria a largo plazo fue almacenada por cambios físicos en la sinapsis ha sido tan influyente que la investigación sobre la base neuronal del aprendizaje y la memoria se ha centrado principalmente en eventos sinápticos (ver Capítulo 5 sobre sinapsis y transmisión sináptica). El enfoque en la sinapsis se vio reforzado por el descubrimiento en 1973 en el hipocampo de conejo de que un aumento duradero en la conductividad sináptica (fuerza sináptica) podría ser producido por la estimulación de alta frecuencia de la neurona presináptica (Bliss y Lomo, 1973). Este hallazgo de que la fuerza sináptica puede aumentarse durante un período prolongado de tiempo después de la estimulación presináptica de alta frecuencia ahora se conoce como Potenciación a Largo Plazo (LTP). La LTP se ha encontrado en muchas especies y en muchas partes del cerebro. Sin embargo, se ha estudiado más en el hipocampo de la rata. Desde esta investigación temprana, también se han descubierto formas adicionales de cambio sináptico.
Plasticidad sináptica
La plasticidad sináptica es el fortalecimiento o debilitamiento de las sinapsis a lo largo del tiempo en respuesta a aumentos o disminuciones en su actividad. El cambio plástico también resulta de la alteración del número de receptores localizados en una sinapsis. La plasticidad sináptica es la base del aprendizaje y la memoria, permitiendo un sistema nervioso flexible y funcional. La plasticidad sináptica puede ser a corto plazo (mejora sináptica o depresión sináptica) o a largo plazo. Dos procesos en particular, la potenciación a largo plazo (LTP) y la depresión a largo plazo (LTD), son formas importantes de plasticidad sináptica que ocurren en las sinapsis del hipocampo.
Plasticidad sináptica: potenciación a corto plazo, potenciación a largo plazo y depresión a largo plazo
Puntos Clave
- El realce sináptico a corto plazo ocurre cuando se incrementa la cantidad de neurotransmisor disponible, mientras que la depresión sináptica a corto plazo ocurre cuando disminuye la cantidad de vesículas con neurotransmisores.
- Las sinapsis se fortalecen en la potenciación a largo plazo (LTP) cuando se incrementan los receptores AMPA (que se unen al glutamato cargado negativamente), permitiendo que más iones calcio ingresen a la célula, provocando una mayor respuesta excitadora.
- La depresión a largo plazo (LTD) ocurre cuando se disminuyen los receptores AMPA, lo que disminuye la cantidad de iones de calcio que ingresan a la célula, debilitando la respuesta sináptica a la liberación de neurotransmisores.
- El fortalecimiento y debilitamiento de las sinapsis a lo largo del tiempo controla el aprendizaje y la memoria en el cerebro.
Términos Clave
- Potenciación a largo plazo: un incremento de larga duración (horas in vitro, semanas a meses in vivo), típicamente en amplitud, de la respuesta de una neurona postsináptica a un patrón de estímulos de alta frecuencia de una neurona presináptica
- depresión a largo plazo: un debilitamiento a largo plazo de una conexión sináptica
- plasticidad: la propiedad de la conductividad o fuerza sináptica de la neurona que permite fortalecerla o debilitarla
- Receptor NMDA: receptor postsináptico de N-metil-D-aspartato (NMDA) y canal iónico que se activa cuando el transmisor de glutamato se une a él; un receptor ionotrópico (ver Capítulo 5) de glutamato
- Receptor AMPA: receptor post-sináptico de ácido alfa-amino-3-hidroxi-5-metil-4-isoxazol propiónico (AMPA) y canal iónico que se activa cuando el transmisor de glutamato se une a él; un receptor ionotrópico (ver Capítulo 5) de glutamato; tanto los receptores NMDA como los receptores AMPA son importantes en el aprendizaje y la memoria
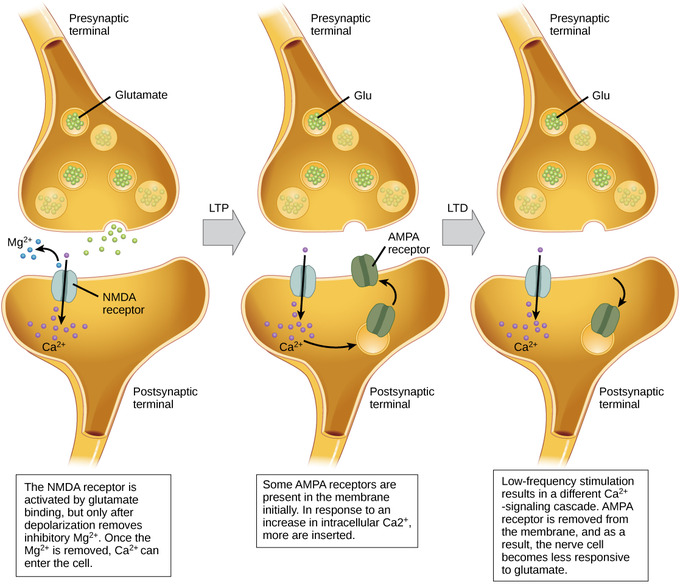
Figura\(\PageIndex{4}\): Potenciación y depresión a largo plazo: La entrada de calcio a través de receptores postsinápticos de NMDA puede iniciar dos formas diferentes de plasticidad sináptica: la potenciación a largo plazo (LTP) y la depresión a largo plazo (LTD). La LTP surge cuando una sola sinapsis es estimulada repetidamente. Esta estimulación provoca una cascada celular dependiente de calcio y CaMKII, lo que resulta en la inserción de más receptores AMPA en la membrana postsináptica. La próxima vez que se libere glutamato de la célula presináptica, se unirá tanto a NMDA como a los receptores AMPA recién insertados, despolarizando así la membrana de manera más eficiente. LTD ocurre cuando pocas moléculas de glutamato se unen a receptores de NMDA en una sinapsis (debido a una baja velocidad de disparo de la neurona presináptica). El calcio que sí fluye a través de los receptores NMDA inicia una cascada diferente dependiente de calcineurina y proteína fosfatasa 1, lo que resulta en la endocitosis de los receptores AMPA. Esto hace que la neurona postsináptica sea menos sensible al glutamato liberado de la neurona presináptica. (Imagen y subtítulo de Lumen Boundless Biology, How Neurons Comunicate; https://courses.lumenlearning.com/bo...s-communicate/; curación y revisión. Proporcionado por: Boundless.com. Licencia: CC BY-SA: Atribución-CompartirIgual. )
Potenciación a largo plazo (LTP)
La potenciación a largo plazo (LTP) es un fortalecimiento persistente de una conexión sináptica, que puede durar minutos u horas o incluso semanas. LTP se basa en el principio hebbio: “las células que disparan juntas se conectan por cable”. Existen diversos mecanismos, ninguno de los cuales se entiende completamente, detrás del fortalecimiento sináptico visto con LTP.
Un mecanismo conocido involucra un tipo de receptor postsináptico de glutamato: los receptores NMDA (N-metil-D-aspartato). Estos receptores normalmente están bloqueados por iones de magnesio. Sin embargo, cuando la neurona postsináptica es despolarizada por múltiples entradas presinápticas en rápida sucesión (ya sea de una neurona o múltiples neuronas), los iones de magnesio son expulsados y los iones Ca 2 + pasan a la célula postsináptica. A continuación, los iones Ca 2 + que ingresan a la célula inician una cascada de señalización que provoca que un tipo diferente de receptor de glutamato, los receptores AMPA (ácido α-amino-3-hidroxi-5-metil-4-isoxazolpropiónico), se inserten en la membrana postsináptica. Los receptores AMPA activados permiten que los iones positivos ingresen a la célula.
Por lo tanto, la próxima vez que se libere glutamato de la membrana presináptica, tendrá un mayor efecto excitatorio (EPSP) sobre la célula postsináptica debido a que la unión del glutamato a estos receptores AMPA permitirá que entren más iones positivos en la célula. La inserción de receptores AMPA adicionales fortalece la sinapsis de manera que la neurona postsináptica es más probable que se active en respuesta a la liberación de neurotransmisores presinápticos.
LTP tiene muchas similitudes con los cambios sinápticos que Hebb propuso como base para la memoria a largo plazo, incluyendo dos características clave. Primero, LTP es de larga duración, hasta un año con ensayos repetidos. En segundo lugar, muchas formas de LTP requieren activación simultánea en neuronas presinápticas y possinápticas en la sinapsis donde tiene lugar la LTP (“neuronas que disparan juntas, cableadas”); esto se debe a que los receptores NMDA (N-metil-D-aspartato) para glutamato (el transmisor excitatorio más común en el cerebro), que son prominentes en las sinapsis donde ocurre la LTP, tienen el mismo requisito de actividad simultánea tanto en neuronas pre y possinápticas para que estos receptores se activen. La investigación que muestra que muchas características de LTP son similares a las características de la memoria a largo plazo proporciona una fuerte evidencia circunstancial de que la LTP está relacionada con los mecanismos de aprendizaje y memoria. Por ejemplo, la LTP puede ser estimulada por estimulación de baja intensidad similar a la producida en neuronas individuales; la LTP es más prominente en las estructuras asociadas al aprendizaje; la LTP se produce en el hipocampo mediante el aprendizaje; las drogas que potencian o perjudican el aprendizaje también mejoran o perjudican la LTP; y la LTP ocurre en los nervios sistema de invertebrados simples (ver discusión de Aplysia arriba) en las sinapsis específicas involucradas en el aprendizaje (Pinel & Barnes, 2021).
Depresión a largo plazo (LTD)
Otro fenómeno relacionado es la depresión a largo plazo (LTD), asociada con disminuciones en la conductividad sináptica. LTD podría ser parte de los procesos involucrados en la creación y modificación de patrones de excitación e inhibición en grandes poblaciones de neuronas para la codificación de movimientos aprendidos, experiencia sensorial, y tal vez las representaciones mentales de estructuras cognitivas complejas como percepciones, memorias enteras, conceptos, e incluso ideas abstractas (Churchland, 2013). La codificación de eventos mentales en términos de patrones de excitación e inhibición en grandes poblaciones de neuronas se discute más adelante en este capítulo cuando examinamos el modelado de redes neuronales del aprendizaje y la memoria. Una posibilidad relacionada es la poda de algunas sinapsis para formar cambios permanentes en el circuito neural.La depresión a largo plazo (LTD) es esencialmente la inversa de LTP: es un debilitamiento a largo plazo de una conexión sináptica. Un mecanismo que se sabe que causa el LTD también involucra a los receptores AMPA. En esta situación, el calcio que ingresa a través de los receptores NMDA inicia una cascada de señalización diferente, lo que resulta en la eliminación de los receptores AMPA de la membrana postsináptica. Con la disminución de los receptores AMPA en la membrana, la neurona postsináptica responde menos al glutamato liberado de la neurona presináptica. Si bien puede parecer contradictorio, LTD puede ser tan importante para el aprendizaje y la memoria como LTP. El debilitamiento y poda de sinapsis no utilizadas recorta conexiones poco importantes, dejando solo las conexiones salientes fortalecidas por la potenciación a largo plazo.
Mejora sináptica a corto plazo y depresión
La plasticidad sináptica a corto plazo actúa en una escala de tiempo de decenas de milisegundos a pocos minutos. La mejora sináptica a corto plazo resulta de más terminales sinápticos que liberan transmisores en respuesta a potenciales de acción presináptica. Las sinapsis se fortalecerán por un corto tiempo debido a un aumento en el tamaño del grupo fácilmente liberable de transmisores empaquetados o un aumento en la cantidad de transmisor empaquetado liberado en respuesta a cada potencial de acción. El agotamiento de estas vesículas fácilmente liberables causa fatiga sináptica. La depresión sináptica a corto plazo también puede surgir de procesos possinápticos y de la activación por retroalimentación de receptores presinápticos.
Cambios en la sinapsis correlacionados con el aprendizaje y la memoria: cambios anatómicos en espinas dendríticas
Las modificaciones en las fuerzas sinápticas en sinapsis excitatorias en el hipocampo parecen desempeñar un papel crítico en el almacenamiento y recuerdo de información en mamíferos. Al menos en humanos, el papel del hipocampo es especialmente crítico en la memoria episódica explícita (autobiográfica). Como se señaló anteriormente, también está involucrado de manera crucial en la memoria espacial de muchas especies de mamíferos, incluidos los humanos. Los cambios en la fuerza sináptica en gran parte del cerebro pueden ser importantes en el aprendizaje y la memoria.
Espinas dendríticas
Las espinas dendríticas son pequeñas estructuras sobresalientes ubicadas en el eje de las dendritas y están asociadas con conexiones sinápticas. Estas espinas “están presentes en grandes cantidades en la superficie de las dendritas. Por ejemplo, una única neurona piramidal en la región CA1 del hipocampo posee hasta 30,000 espinas dendríticas. La mayoría de las sinapsis excitatorias se forman en la superficie de estas espinas dendríticas” (Irie & Yamaguchi, 2009, p. 1141). Los sitios primarios de interacción sináptica excitatoria en el sistema nervioso central de los mamíferos parecen estar en las espinas dendríticas.
Estas protuberancias espinosas tienen cabeza y cuello de morfología variada (ver Figuras 10.4.5 y 10.4.6). Los cambios en las espinas juegan un papel importante durante el desarrollo cerebral. “En un cerebro en desarrollo, las espinas exhiben un alto grado de plasticidad estructural y funcional, reflejando la formación y eliminación de sinapsis durante la maduración de los circuitos neuronales. La morfología de las espinas en las neuronas en desarrollo se ve afectada por la actividad sináptica, contribuyendo así al refinamiento dependiente de la experiencia de los circuitos neuronales, el aprendizaje y la memoria. Así, comprender la dinámica de la columna vertebral y su regulación es de importancia central para los estudios de plasticidad sináptica en el cerebro” (Bertling, et al., 2012, p. 391). Durante el desarrollo cerebral se forman sinapsis, se modifican y en ocasiones se eliminan en función de la entrada a las mismas.
Como se discutió anteriormente, se ha demostrado que la LTP conduce a cambios funcionales en las sinapsis donde se induce. Estos cambios debidos a la LTP son una mayor conductividad sináptica y una mayor capacidad de respuesta de la neurona possináptica. Parece que los cambios anatómicos en las espinas acompañan a estos cambios en la fuerza sináptica que ocurren durante la LTP y el aprendizaje y la memoria.
Recuerde la propuesta de Hebb de que el almacenamiento de memoria a largo plazo implica algún cambio anatómico persistente en las sinapsis. Si Hebb tenía razón, entonces deberíamos esperar ver cambios anatómicos en las sinapsis cuando ocurren el aprendizaje y la memoria. Y si la LTP está involucrada en la formación de estos cambios anatómicos hipotéticos, entonces deberíamos esperar ver modificaciones anatómicas en la sinapsis asociadas con los aumentos inducidos por LTP en la conductividad sináptica y la capacidad de respuesta de la neurona possináptica. Consistente con la hipótesis de Hebb sobre la base física del aprendizaje y la memoria, esto es justo lo que los neurocientíficos han observado. Los cambios en la fuerza sináptica, asociados a LTP, van acompañados de alteraciones inducidas por LTP de la forma y tamaño de las espinas dendríticas (Chidambaram, et al., 2019; Harris, et al., 2003), cambios anatómicos en la sinapsis, tal como predijo Hebb.
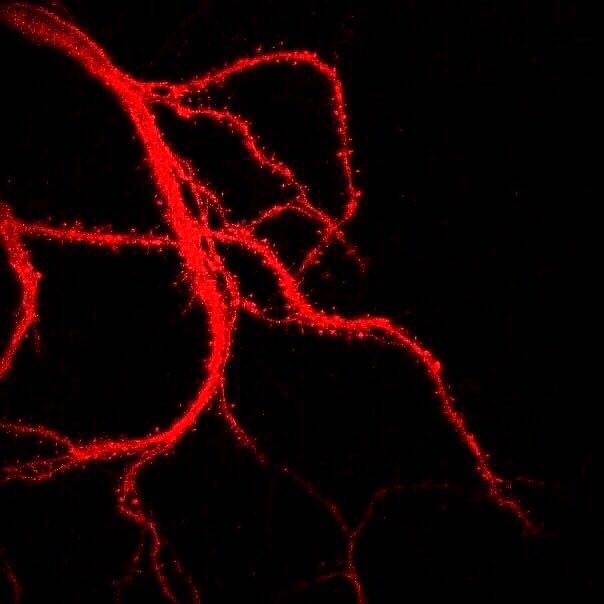
Figura\(\PageIndex{5}\): Dendrítas ramificadas de una neurona que muestran espinas dendríticas (pequeñas proyecciones parecidas a cerdas que recubren cada rama dendrítica). La fotografía se obtuvo con un microscopio de barrido láser. Las espinas dendríticas pueden cambiar rápidamente en tamaño, forma y números, y son importantes en el aprendizaje y la memoria. (Leyenda de Kenneth A. Koenigshofer, PhD. Imagen de Wikimedia Commons; Archivo:Нейрональные отростки с шипиками.jpg; https://commons.wikimedia.org/wiki/F...0%BC%D0%B8.jpg; por Sergb95; con licencia Creative Commons Attribution-Share Alike 4.0 International).
La evidencia sugiere fuertemente que la nueva síntesis de proteínas es necesaria para estos efectos sobre la forma y el tamaño de las espinas dendríticas (Chidambaram, et al., 2019). Como se señaló anteriormente, tanto el crecimiento como los cambios metabólicos en las sinapsis fueron propuestos por Hebb como posible base para la memoria a largo plazo. Estos cambios anatómicos en las espinas dendríticas inducidos por LTP pueden ser duraderos. Por ejemplo, “En redes maduras, las conexiones sinápticas en las espinas dendríticas pueden ser bastante estables, ya que se ha demostrado que las espinas recién emergentes generadas después del aprendizaje motor persisten durante meses. Si bien la distribución del tamaño de la columna vertebral a través del árbol dendrítico de una sola neurona puede ser bastante variable, el tamaño de la columna generalmente se correlaciona con la fuerza de la sinapsis excitatoria. Generalmente se acepta que el diámetro de la cabeza de la columna y la fuerza de sinapsis covarían durante la expresión de la potenciación a largo plazo (LTP). .” (Henry, et al., 2017). Parece que la hipótesis de Hebb sobre la base física del aprendizaje y la memoria en el cerebro era bastante predictiva de lo que mostraría la investigación posterior, una marca de una buena teoría científica.
De acuerdo con la hipótesis de Hebb, las espinas dendríticas parecen funcionar como sitios de almacenamiento para la fuerza sináptica y ayudan a transmitir potenciales eléctricos al cuerpo celular de la neurona. Las dendritas de una sola neurona pueden contener cientos, incluso miles de espinas. Las espinas dendríticas suelen recibir entrada excitatoria de los axones, aunque a veces se realizan conexiones tanto inhibidoras como excitadoras sobre la misma espina dendrítica (Kasthuri, et al., 2015). Además de que las espinas proporcionan un sustrato anatómico potencial para el almacenamiento de la memoria y la transmisión sináptica, también pueden aumentar el número de posibles contactos entre neuronas (Alvarez & Sabatini, 2007). Las espinas se encuentran en las dendritas de la mayoría de las neuronas principales del cerebro, incluyendo las neuronas piramidales del neocórtex (en la corteza prefrontal para la cognición; tracto corticoespinal para el movimiento), las neuronas espinosas medianas (inhibidoras GABAérgicas) del cuerpo estriado (núcleo estriado de los ganglios basales; motor y recompensa ; recibe entradas glutamatérgicas y dopaminérgicas), y las células de Purkinje (neuronas inhibidoras GABAérgicas) del cerebelo que reciben sinapsis excitadoras (glutamatérgicas) relativamente más débiles a las espinas en la dendrita celular de Purkinje. Las neuronas piramidales del hipocampo y corticales pueden recibir decenas de miles de entradas en su mayoría excitatorias de otras neuronas en sus espinas, mientras que el número de espinas en las dendritas de neuronas de Purkinje en el cerebelo es aún mayor, hasta diez veces mayor.
Tres de las cuatro clases más notables de forma de columna vertebral se muestran en la siguiente figura: “delgado”, “rechoncho” y “hongo”. No se muestra la cuarta categoría, “ramificada”. Estudios con microscopía electrónica han revelado un continuo de formas entre estas categorías (Ofer, et al., 2021). La variable forma y volumen de la columna vertebral se correlaciona con la fuerza y madurez de cada sinapsis espinal. “Las espinas dendríticas también son altamente móviles y experimentan cambios de tamaño y forma a lo largo de una escala de tiempo de segundos a minutos. Debido a que las espinas dendríticas pequeñas y delgadas tienen más probabilidades de sufrir estos cambios estructurales, mientras que las espinas grandes, las llamadas “setas” tienden a mantener su forma, se ha sugerido que las espinas de los hongos son espinas de memoria más estables mientras que las espinas delgadas más plásticas son espinas de aprendizaje. Las espinas [L] arge son sitios de sinapsis fuertes y, en consecuencia, el crecimiento de la cabeza de la columna probablemente se correlaciona con un fortalecimiento de la transmisión sináptica” (Leuner & Shors, 2010, p. 349).
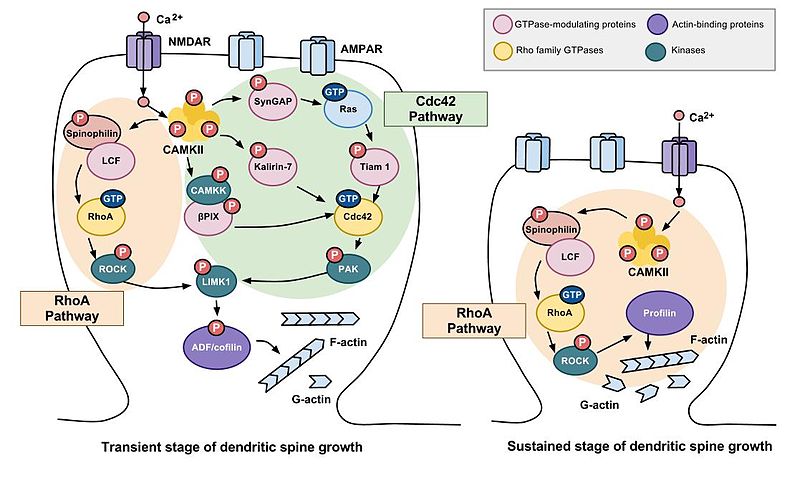
Los investigadores han observado cambios tanto transitorios a corto plazo como sostenidos a largo plazo en la plasticidad estructural de las espinas dendríticas; dentro de los dos primeros minutos siguientes a la estimulación de la neurona presináptica, se produce una expansión inicial del 300% de la columna dendrítica postsináptica, seguida de una reducción del volumen de la columna vertebral para que se eleve a aproximadamente 70-80% más grande que el volumen original de pre-estimulación, un cambio sostenido en la plasticidad estructural que dura aproximadamente 30 minutos o más. La LTP se inicia con la etapa transitoria del crecimiento de la columna dendrítica. Se plantea la hipótesis de que ambas etapas del crecimiento de la columna están involucradas en el aprendizaje y la memoria. Las espinas grandes son más estables que las más pequeñas y pueden ser resistentes a la modificación por actividad sináptica adicional (Kasai, et al., 2003). Estos cambios estructurales en las espinas dendríticas inducidos por la estimulación presináptica asociada con LTP son considerados por las neurociencias como importantes en la codificación, almacenamiento y recuperación de memorias (Murakoshi, et al., 2011). Estos cambios en el tamaño y forma de la columna implican cambios en la microestructura de la columna y anomalías a este nivel pueden estar involucradas en trastornos de la memoria. “El citoesqueleto de actina es el elemento estructural que subyace a los cambios en la morfología de la columna dendrítica y la fuerza Se ha demostrado que la morfología adecuada de las espinas y la regulación adecuada del citoesqueleto de actina son importantes en la memoria y el aprendizaje; los defectos en la regulación conducen a diversos trastornos de la memoria. Así, entender la regulación del citoesqueleto de actina en espinas dendríticas es de importancia central para los estudios de la función sináptica y neuronal” (Koskinen, et al., 2012, p. 47).
La morfogénesis de espinas dendríticas (proceso por el cual alcanzan su forma) es crítica para la inducción de potenciación a largo plazo (LTP) (Kim & Lisman, 1999; Krucker, et al., 2000). Curiosamente, Hayashi-Takagi et al. (2015) encontraron que la memoria podría alterarse si las espinas potenciadas, dentro de un conjunto de neuronas involucradas en el aprendizaje motor, se redujeran específicamente. Esto sugiere que el crecimiento de la columna vertebral es esencial para al menos algunas formas de memoria.

Figura\(\PageIndex{6}\): (Izquierda) Morfología variable de tipos de espinas dendríticas. Los cambios en su forma y tamaño están asociados con LTP y aprendizaje y memoria. (Derecha) Espinas en la dendrita de una neurona estriatal media espinosa. La imagen se obtuvo mediante la expresión de Proteína Verde Fluorescente Mejorada (EGFP) en las neuronas y la obtención de imágenes mediante un microscopio de dos fotones de barrido láser. (Leyenda para imagen a la izquierda por Kenneth A. Koenigshofer, PhD; Imagen a la izquierda de Wikimedia Commons, Espinas dendríticas; https://commons.wikimedia.org/wiki/F...e_types_3D.png; obra original de Thomas Splettstoesser (www.scistyle.com); licenciada bajo la Creative Commons Reconocimiento-Compartir Igual 3.0 Licencia Unported. Imagen a la derecha y su leyenda de Wikimedia Commons, Espinas dendríticas; https://commons.wikimedia.org/w/inde... =go&type=image; Liberado al dominio público por el autor de la imagen).
Figura\(\PageIndex{7}\): Crecimiento de la Columna Dendrítica Transitoria vs Sostenida después de Estimulación Los detalles dentro de cada columna muestran las cascadas químicas involucradas en las dos etapas de crecimiento de la columna después de la estimulación presináptica. No es necesario que el alumno conozca los detalles de estas cascadas. Sin embargo, tenga en cuenta que estas cascadas comienzan con la afluencia de calcio a través de los receptores NMDA (parte superior de la columna vertebral) y que el paso final es la acción sobre la actina (parte inferior derecha de la columna) que conduce a cambios en la forma y el tamaño de la columna vertebral. (Imagen de Wikimedia Commons; Espinas dendríticas; https://commons.wikimedia.org/wiki/F...timulation.jpg; autor de la imagen y título de la imagen, Itzy; licenciado bajo la licencia Creative Commons Reconocimiento-Compartir Igual 3.0 Unported; caption by Kenneth A. Koenigshofer, PhD.).
Yang, et al. (2009) estudiaron los cambios en el número de espinas sinápticas durante el aprendizaje y la experiencia sensorial novedosa en ratones. Afirman que sus “resultados sugieren que una nueva experiencia conduce a la poda de sinapsis existentes y podría provocar cambios funcionales significativos en los circuitos corticales. De hecho, encontramos que 1 semana después del entrenamiento motor, el rendimiento motor se correlacionó fuertemente con el grado de eliminación de la columna vertebral. Así, el aprendizaje motor y la experiencia sensorial novedosa implican no solo la formación de nueva columna sino también la eliminación permanente de conexiones establecidas temprano en la vida” (Yang, et al. 2009, p. 921). Estos resultados son más fáciles de entender si recordamos que durante el desarrollo prenatal del cerebro hay un número excesivo de conexiones sinápticas y que la eliminación de las sinapsis no funcionales es esencial para el desarrollo y funcionamiento normal del cerebro. El aprendizaje puede implicar poda neural, así como la formación y mejora de la conductividad de las sinapsis a través de cambios en las espinas dendríticas. Como Yang, et al., señalan, “Además de promover la formación de sinapsis, la experiencia juega un papel importante en la eliminación de conexiones sinápticas excesivas e imprecisas formadas temprano durante el desarrollo” (Yang, et al. 2009, p. 921).
Chidambaram, et al. (2019, p.161) resumen la relación de los cambios en la conductividad sináptica con los cambios en las espinas dendríticas: “Durante la plasticidad sináptica el número y las formas de las espinas dendríticas experimentan reorganizaciones radicales. La inducción de potenciación a largo plazo (LTP) promueve el agrandamiento de la columna vertebral y la formación y estabilización de nuevas espinas. La depresión a largo plazo (LTD) resulta en su contracción y retracción”. Estos autores también señalan algunas observaciones interesantes que relacionan números atípicos de espinas sinápticas con varios trastornos de base neurológica. “Los informes indican aumento de la densidad de la columna vertebral en las neuronas piramidales de pacientes con autismo y síndrome del X frágil y densidad reducida en los loci de circunvolución temporal de pacientes esquizofrénicos. Los reportes post mortem de cerebros de Alzheimer mostraron un número reducido de espinas en el hipocampo y la corteza” y las espinas atípicas pueden jugar un papel en las enfermedades neurodegenerativas (Chidambaram, et al., 2019, p. 161).
Si los cambios en las espinas están involucrados en la formación y retención de recuerdos a largo plazo, entonces los cambios morfológicos en las espinas deben ser muy duraderos. Según Yang y sus colegas (2009, p. 920), “Las espinas dendríticas mantenidas de manera estable están asociadas con recuerdos de por vida.... una pequeña fracción de nuevas espinas inducidas por la experiencia novedosa, junto con la mayoría de las espinas formadas temprano durante el desarrollo y supervivientes a la eliminación dependiente de la experiencia, se conservan a lo largo de toda la vida de un animal. Estos estudios indican que el aprendizaje y la experiencia sensorial diaria dejan marcas minuciosas pero permanentes en las conexiones corticales y sugieren que los recuerdos de toda la vida se almacenan en redes sinápticas conectadas en gran parte de manera estable”.
La teoría de Hebb ha demostrado ser impresionantemente predictiva de los descubrimientos realizados muchos años después de que su libro fuera publicado en 1949. Hallazgos como los descritos en este módulo continúan confirmando muchos aspectos de la teoría de Hebb.
Mecanismos de Nivel Celular y Receptor Revisitados
En los apartados anteriores, se discutió la Potenciación a Largo Plazo (LTP) y su posible implicación en los procesos de memoria. Aquí, es apropiado considerar la LTP y los procesos asociados con mayor detalle. Como se señaló anteriormente, la potenciación a largo plazo (LTP) es un proceso en el que se fortalecen las sinapsis. Ha sido objeto de una extensa investigación desde mediados de la década de 1970 debido a su probable papel en varios tipos de memoria.
Un fenómeno relacionado que también se discutió anteriormente es la Depresión a Largo Plazo (LTD), un fenómeno que es lo opuesto a la potenciación a largo plazo (LTP). En LTD, la comunicación a través de la sinapsis es silenciada. LTD juega un papel importante en el cerebelo, en la memoria procedimental implícita (memoria motora, memoria implícita de cómo hacer algo como andar en bicicleta o golpear una pelota de béisbol), donde las redes neuronales involucradas en movimientos erróneos son inhibidas por el silenciamiento de sus conexiones sinápticas. LTD es lo que nos permite corregir nuestros procedimientos motores a la hora de aprender a realizar una tarea. Así, la depresión a largo plazo (LTD) implica un debilitamiento de una sinapsis como medio para mejorar el rendimiento de los movimientos expertos aprendidos (recordemos que el debilitamiento de la fuerza sináptica se asocia con la contracción de las espinas dendríticas). Imagina que estás tratando de aprender una rutina de gimnasia que implica una serie de volteretas y otros movimientos complicados en secuencia. Cuando inicialmente comienzas a aprender la rutina, haces muchos movimientos incorrectos que debes suprimir si vas a aprender a hacer la rutina correctamente con la precisión libre de errores que buscarán los jueces. LTD es el mecanismo a nivel celular que permite suprimir los movimientos musculares incorrectos que conducen a errores en la rutina.
Como se discutió anteriormente, existe otro posible papel para el LTD en la codificación de percepciones, conceptos, ideas y otras estructuras cognitivas complejas. Uno de los problemas más difíciles de la neurociencia es cómo las actividades de grandes poblaciones de neuronas codifican y crean elementos complejos de la experiencia psicológica como percepciones, pensamientos, ideas, comprensión, conocimiento y otras representaciones mentales complejas. El modelado de procesos cerebrales en redes neuronales artificiales (ver el suplemento más adelante en este capítulo sobre redes artificiales) sugiere que la inhibición es tan importante como la excitación en la codificación neuronal de representaciones mentales complejas. Los patrones de excitación e inhibición en grandes poblaciones de neuronas pueden ser la base de la codificación neural de la cognición y percepción complejas. Si es así, entonces LTD puede ser igual de importante que LTP en la codificación neuronal de representaciones complejas como percepciones, pensamientos, conceptos, ideas e incluso emociones (ver Materia y conciencia de Paul Churchland para una discusión más completa de estas ideas). Así como los puntos y guiones son igualmente importantes en el Código Morse, la excitación y la inhibición pueden ser elementos igualmente importantes en la codificación neuronal.
Aunque una sinapsis se debilita en el LTP, por el contrario, en LTP, después de una intensa estimulación de la neurona presináptica, se fortalece una sinapsis y aumenta la amplitud de la respuesta de la neurona possináptica. El estímulo aplicado a la neurona presináptica para producir LTP es generalmente de corta duración (menos de 1 segundo) pero de alta frecuencia (más de 100 Hz). El potencial excitatorio (EPSP; ver Capítulo 5) medido en una neurona possináptica, después de la estimulación presináptica de alta frecuencia, se incrementa por un largo periodo. Por ejemplo, cuando los axones que hacen conexiones a las neuronas piramidales del hipocampo se exponen a un estímulo de alta frecuencia, la amplitud del potencial excitatorio medido en estas neuronas piramidales se incrementa hasta por varias semanas. ¿Qué eventos sinápticos están involucrados en la producción de LTP? Repasemos, ahora con mayor detalle.
El glutamato, el neurotransmisor liberado en estas sinapsis del hipocampo (el glutamato es excitatorio y media la neurotransmisión sináptica rápida en el cerebro), se une a varios subtipos diferentes de receptores en la neurona hipocampal possináptica. Dos de estos subtipos de receptores de glutamato, los receptores para AMPA y NMDA, son especialmente importantes para LTP (Traynelis, et al., 2010).
El receptor AMPA (un receptor de glutamato que lleva el nombre del derivado que lo activa, AMPA abreviatura de Ácido Alfa-Amino-3-Hidroxi-5-Metil-4-Isoxazol Propiónico) se empareja con un canal iónico para que cuando el glutamato se une a este receptor, este canal permita la entrada de iones de sodio la neurona possináptica. Esta afluencia de sodio hace que la dendrita possináptica se despolarice localmente (un EPSP, un cambio positivo en el voltaje).
El receptor NMDA, que también utiliza glutamato como transmisor, también está emparejado con un canal iónico (NMDA es N-metil-D-aspartato, un derivado del glutamato). Este canal admite iones de calcio en la célula possináptica cuando se activa. Sin embargo, cuando la célula está en potencial de reposo, el canal de calcio es bloqueado por iones magnesio (Mg2+), de manera que incluso si el glutamato se une al receptor, el calcio no puede ingresar a la neurona. Para que estos iones de magnesio se retiren del canal, se debe despolarizar el potencial de membrana de la dendrita. Y eso es exactamente lo que sucede durante la estimulación de alta frecuencia que causa LTP: ¡la neurona possináptica se despolariza tras la activación sostenida de sus receptores AMPA! El magnesio luego se retira de los receptores NMDA y permite que un gran número de iones de calcio ingresen a la célula.
Esta mayor concentración de calcio en la dendrita desencadena varias reacciones bioquímicas que hacen que esta sinapsis sea más eficiente por un período prolongado (Bliss & Collingridge, 1993; Bliss, et al., 2018; Citri & Malenka, 2008). Estos iones de calcio son mensajeros intracelulares extremadamente importantes que activan muchas enzimas alterando su conformación. Una de estas enzimas es la calmodulina, que se activa cuando cuatro iones de calcio se unen a ella. Luego se convierte en Ca2+/calmodulina, el segundo mensajero principal para LTP. Ca2+/calmodulina activa entonces a su vez otras enzimas que desempeñan un papel clave en este proceso, como la adenilato ciclasa y la proteína quinasa II dependiente de Ca2+/calmodulina (CaM quinasa II). Estas enzimas a su vez modifican la conformación espacial de otras moléculas, generalmente agregándoles un ion fosfato. Este proceso catalítico común se llama fosforilación.
La adenilato ciclasa activada fabrica monofosfato de adenosina cíclico (AMPc), que a su vez cataliza la actividad de otra proteína, la quinasa A (PKA). En definitiva, existe una cascada típica de reacciones bioquímicas que pueden tener muchos efectos diferentes.
Por ejemplo, PKA fosforila los receptores AMPA, permitiendo que permanezcan abiertos más tiempo después de que el glutamato se una a ellos. Como resultado, la neurona possináptica se despolariza aún más, contribuyendo así a la LTP.
Otros experimentos han demostrado que la proteína CREB es otra diana de PKA, la proteína quinasa A. CREB juega un papel importante en la transcripción génica, y su activación conduce a la creación de nuevos receptores AMPA que pueden aumentar aún más la eficiencia sináptica.
Según Park, et al. (2021), la potenciación a largo plazo (LTP) en las sinapsis CA1 del hipocampo (ver Figura 10.4.8) puede expresarse por un aumento en el número (N) de receptores AMPA (ácido α-amino-3-hidroxi-5-metil-4-isoxazol propiónico) o en su conductancia de canal único (γ), es decir, aumento sináptico fuerza. En sus experimentos, establecieron cómo estos distintos procesos sinápticos contribuyen a la expresión de LTP en rodajas de hipocampo obtenidas de roedores adultos jóvenes. La LTP inducida por estimulación de estallido theta comprimido (TBS), con un intervalo entre episodios de 10 segundos, implica puramente un aumento en el número de receptores AMPA (LTP N). En contraste, ya sea un TBS espaciado, con un intervalo entre episodios de 10 min, o un solo TBS, entregado cuando se activa PKA, da como resultado LTP que se asocia con un aumento transitorio en la conductancia de canal único (γ) o una mayor fuerza sináptica (LTP γ), causada por la inserción de permeables al calcio Receptores (CP) -AMPA. La activación de CaMKII es necesaria y suficiente para LTP N (aumento en el número de receptores AMPA, mientras que PKA se requiere adicionalmente para LTP γ (aumento en la conductancia de canal único). Así, dos formas mecanísticamente distintas de LTP coexisten en estas sinapsis.
La otra enzima activada por Ca2+/calmodulina, CaM quinasa II, tiene una propiedad que es decisiva para la persistencia de LTP: ¡puede fosforilarse a sí misma! Su actividad enzimática continúa mucho después de que el calcio haya sido evacuado de la célula y se haya desactivado el Ca2+/calmodulina.
La CaM quinasa II puede entonces a su vez fosforilar los receptores AMPA y probablemente otras proteínas como las MAP quinasas, que están involucradas en la construcción de dendritas, o los propios receptores NMDA, cuya conductancia de calcio se vería incrementada por esta fosforilación.
| La LTP implica al menos dos fases: establecimiento (o inducción), que dura aproximadamente una hora, y mantenimiento (o expresión), que puede persistir por varios días. | La primera fase, el establecimiento, puede ser inducida experimentalmente por una única estimulación de alta frecuencia. Implica la actividad de diversas enzimas (quinasas) que persisten después de eliminar el calcio, pero no se produce síntesis de proteínas. | Para desencadenar la fase de mantenimiento, sin embargo, se debe aplicar una serie de estímulos de alta frecuencia. A diferencia de la fase de establecimiento de LTP, la fase de mantenimiento requiere la síntesis de nuevas proteínas, por ejemplo, las que forman los receptores y las que contribuyen al crecimiento de nuevas sinapsis (otro fenómeno que ocurre durante la fase de mantenimiento de LTP). |
Cuadro 10.4.1: Fases de Establecimiento y Mantenimiento de LTP
Para que el calcio entre en la célula, el receptor NMDA debe ser activado por glutamato y sometido a despolarización simultáneamente. La necesidad de estas dos condiciones simultáneas le da a este receptor propiedades asociativas. Esto le permite detectar la coincidencia de dos eventos y lo convierte en el elemento clave en la potenciación a largo plazo.
Significativamente, si este receptor está bloqueado con un fármaco, o si el gen involucrado en su construcción está deshabilitado, la LTP no puede ocurrir.
Las espinas en las dendritas possinápticas forman compartimentos separados para aislar las reacciones bioquímicas que ocurren en algunas sinapsis pero no en otras. Esta especialización anatómica probablemente ayude a asegurar una cierta especificidad en las conexiones neuronales.
La característica más interesante de la LTP es que causa el fortalecimiento a largo plazo de las sinapsis entre dos neuronas que se activan simultáneamente. En otras palabras, exactamente el tipo de mecanismo de asociación que Hebb (1949) había imaginado años antes.
Evidencia Experimental para Componentes de LTP
Se han utilizado muchos métodos para determinar el papel de un ion particular, o un segundo mensajero, o una enzima en un proceso sináptico.
Por ejemplo, el papel del calcio en la potenciación a largo plazo (LTP) se ha confirmado de varias maneras. Un experimento consistió en inyectar a la neurona possináptica agentes quelantes como EGTA y BAPTA, dos moléculas que se unen al calcio y lo vuelven inactivas. Como resultado, se vuelve imposible inducir LTP. También se ha utilizado el procedimiento inverso. Los investigadores han inyectado moléculas especiales en la neurona possináptica y luego las iluminaron con luz UV, provocando así que liberen suficiente calcio para inducir LTP en esta neurona.
Otro enfoque es producir mutaciones que hagan que una proteína no sea funcional o que bloqueen su acción con otra molécula. El bloqueo de CaM quinasa II de esta manera evita que la LTP se establezca, lo que también demuestra el papel central de esta proteína.
De igual manera, la inhibición de PKA o CREB impide que LTP alcance su segunda fase y se mantenga.
En ciertos casos, los investigadores incluso han identificado el único aminoácido, entre los cientos de aminoácidos que componen una proteína, cuya fosforilación es esencial para LTP (por si quieres saber: Ser831 para la subunidad GluR1 del receptor AMPA y Thr286 para el sitio de autofosforilación de CaM quinasa II).
Los investigadores también han demostrado que un ratón cuyo gen para la subunidad GluR1 del receptor AMPA había sido noqueado no podía tener ningún LTP, confirmando así el papel de CaM quinasa II una vez más.
En ratones para los que se desactivó el sitio Thr286 en CaM quinasa II, se mantuvo su transmisión sináptica básica, pero ya no se pudo inducir LTP en ellos, demostrando así la necesidad de CaM quinasa II. El procedimiento inverso también condujo a la misma conclusión: agregar CaM quinasa II activada a las neuronas piramidales del hipocampo provoca un fenómeno de potenciación similar al LTP.
Mecanismos adicionales de LTP
Las “sinapsis silenciosas” son otro mecanismo que se descubrió a mediados de la década de 1990 y que puede contribuir a la potenciación a largo plazo (LTP). Estas sinapsis están físicamente presentes, pero en condiciones normales no contribuyen a la transmisión sináptica.
Algunas de estas sinapsis silenciosas se han encontrado en el hipocampo. Parecen tener receptores para NMDA pero no para AMPA. Se piensa que estas sinapsis pueden activarse durante la LTP y así ayudar a fortalecer la respuesta sináptica. El descubrimiento de que después de la LTP, estas sinapsis muestran una corriente eléctrica asociada con los canales AMPA sugiere que algunos receptores AMPA recién sintetizados pueden insertarse en la membrana possináptica.
Además de todos los mecanismos possinápticos involucrados en el establecimiento de LTP, se ha postulado desde hace tiempo que algunas modificaciones presinápticas ocurren durante la fase de mantenimiento subsiguiente. Pero ciertas modificaciones, como un aumento en la cantidad de glutamato liberado por la neurona presináptica, implicarían la presencia de un mensajero retrógrado que se remonta a esta neurona y la modifica. Debido a que el óxido nítrico (NO) es un gas en su estado natural, y así puede difundirse a través de las membranas celulares, sería un candidato ideal para este papel. Pero su implicación sigue siendo objeto de mucho debate y polémica.
Reseñas en vídeo de LTP and Memory
Para una revisión, eche un vistazo a estos breves videos de los eventos en LTP:
https://www.youtube.com/watch?v=KyQUBukwwO8
LTP, cascadas neuroquímicas y etapas de la memoria
Según Rozensweig (2007), diferentes partes de la cascada neuroquímica, asociadas con el aprendizaje y la memoria, pueden estar relacionadas con diferentes etapas en el procesamiento de la memoria. Bennett, et al. (1964), realizaron un descubrimiento temprano de que la experiencia enriquecida en ratas provoca mayores tasas de síntesis de proteínas y mayores cantidades de proteína en la corteza. Mizumori, et al. (1985), utilizando el inhibidor de la síntesis de proteínas, anisomicina, encontraron que la proteína debe sintetizarse en la corteza poco después del entrenamiento si se va a formar LTM; sin embargo, la memoria a corto plazo (STM) no requirió síntesis de proteínas, hallazgos consistentes con los dos tipos de trazas de memoria que Hebb (1949) habían propuesto: trazas transitorias, lábiles de memoria por un lado y trazas estructurales estables por el otro. Utilizando polluelos, varios investigadores rastrearon una cascada de eventos neuroquímicos desde la estimulación sensorial inicial hasta la síntesis de proteínas y cambios estructurales en el cerebro (Rose, 1992). Rozensweig (2007) resume algunos de estos eventos de la siguiente manera:
“La cascada se inicia cuando la estimulación sensorial activa órganos receptores que estimulan las neuronas aferentes mediante el uso de diversos agentes transmisores sinápticos como la acetilcolina (ACh) y el glutamato. Los inhibidores de la actividad sináptica de ACh, como la escopolamina y la pirenzepina, pueden prevenir la STM al igual que los inhibidores de los receptores de glutamato, incluidos los receptores NMDA y AMPA. La alteración de la regulación de los canales iónicos en la membrana neuronal puede inhibir la formación de STM, como se ve en los efectos del cloruro de lantano en los canales de calcio y de la ouabaína en los canales de sodio y potasio. La inhibición de segundos mensajeros también es amnésica, por ejemplo, inhibición de adenilato ciclasa por forskolina o de diacilglicerol por bradiquinina. Estos segundos mensajeros pueden activar las proteínas quinasas, enzimas que catalizan las adiciones de moléculas de fosfato a las proteínas. Se encontró que dos tipos de proteína quinasas son importantes en la formación, respectivamente, de ITM (una etapa intermedia en la formación de la memoria señalada por Rozenzweig, 2007) o LTM. Los agentes que inhiben las proteínas quinasas de calcio/calmodulina (CaM quinasas) previenen la formación de ITM, mientras que los agentes que no inhiben las CaM quinasas, sino que inhiben la proteína quinasa A (PKA) o la proteína quinasa C (PKC) previenen la formación de LTM (Rosenzweig, et al., 1992; Serrano P.A., et al., 1994).”
Rose (1995) sugirió que en los polluelos una especie de LTM que dura algunas horas (ITM de Rozensweig) implica una primera ola de síntesis de glicoproteínas, mientras que la “verdadera memoria a largo plazo” (LTM) requiere una segunda ola de síntesis de glicoproteínas, ocurriendo aproximadamente 6 horas después del entrenamiento.
Rozensweig (2007) revisa evidencias de que las cascadas neuroquímicas en memoria en el polluelo son similares a las cascadas en formación de LTP en diferentes especies. Afirma:
“La cascada neuroquímica involucrada en la formación de la memoria en el polluelo pronto demostró ser similar a la cascada involucrada en la potenciación a largo plazo en el cerebro de los mamíferos (Colley & Routtenberg, 1993) y en los sistemas nerviosos de los invertebrados (Krasne & Glanzman, 1995). DeZazzo y Tully (1995) compararon STM, ITM y LTM en moscas de la fruta, polluelos y ratas. Tully y sus compañeros de trabajo han demostrado que las tres etapas de la memoria en la mosca de la fruta dependen de tres genes diferentes (Tully et al., 1996).”
Investigaciones más recientes sobre LTP también confirman un proceso de tres etapas en LTP. En una revisión de la literatura sobre LTP, Bliss, et al. (2018) señalan:
“Los marcadores LTP1 y LTP2 equivalen a las formas de LTP que son, respectivamente, independientes y dependientes de la síntesis de proteínas de novo. Estos se denominan frecuentemente LTP de fase temprana y LTP de fase tardía (E-LTP y L-LTP, respectivamente), lo que implica que la síntesis de proteínas no se requiere inicialmente sino que se requiere en etapas posteriores, con el cambio [entre E-LTP y L-LTP] ocurriendo durante un periodo de unas pocas horas”.
Aunque la LTP tiene muchas propiedades que la convierten en un buen candidato para el mecanismo de aprendizaje y memoria en el cerebro, aún se necesita evidencia crítica en el comportamiento de los animales. Sin embargo, los neurocientíficos son optimistas, y por una buena razón. Según Bliss, et al. (2018), “Hoy en día, la LTP se puede estudiar en todos los niveles desde lo puramente molecular hasta lo cognitivo. Si bien todavía faltan pruebas definitivas de que los mecanismos de LTP subsirven el aprendizaje y la memoria en el animal que se comporta, pocos neurocientíficos dudan de que tal prueba eventualmente llegue. Ahora hay evidencia muy fuerte de que un mecanismo similar al LTP media al menos algunos aspectos de la memoria”. Gran parte de este optimismo proviene de numerosos estudios que muestran que las manipulaciones fisiológicas, genéticas o farmacológicas de la LTP (ya sea facilitándola o inhibiéndola) tienen efectos similares en el aprendizaje y la memoria (Bliss, et al., 2018; Rozenzweig, 2007).
Subregiones Anatómicas del Hipocampo y Memoria
Como se mencionó anteriormente, las neuronas piramidales del hipocampo y corticales pueden recibir decenas de miles de entradas en su mayoría excitatorias de otras neuronas en sus espinas dendríticas. Como se muestra a continuación en la Figura\(\PageIndex{8}\), el hipocampo consta de una serie de subregiones. Estas regiones están involucradas en diferentes funciones relacionadas con el aprendizaje y la memoria. “El hipocampo propiamente dicho se define por la circunvolución dentada y Cornu Ammonis (CA). Mientras que la circunvolución dentada contiene la fascia dentata y el hilio, la CA se diferencia anatómica y funcionalmente en distintos subcampos denominados CA1, CA2, CA3 y CA4. [T] el subcampo CA3... con entradas de la circunvolución dentada y la corteza entorinal.. está implicado en la codificación de representaciones espaciales y memorias episódicas. La vía de la fibra musgosa... traduce... señales corticales a a.. código hipocampal, esencial para la formación de la memoria” (Cherubini & Miles, 2015, p.19). Según Yang et al. (2014), “las neuronas piramidales CA3 forman extensas conexiones recurrentes entre sí. Tales conexiones son capaces de aprender a asociar componentes de un patrón de entrada entre sí”. Quizás se requeriría esta capacidad para vincular los componentes de una memoria a un todo unificado.
Un modelo destacado de la organización del hipocampo resalta lo que los neurocientíficos han denominado los “circuitos trisinápticos”, circuitos con tres sinapsis (dentado—CA3—CA1), que según la “hipótesis lamelar” se apilan unos sobre otros a lo largo del cuerpo del hipocampo. Según este modelo, “el hipocampo está organizado como una pila de circuitos trisinápticos paralelos” (Yang et al., 2014, p. 12919).
Sin embargo, investigaciones adicionales sugieren una red orientada perpendicular a los circuitos trisinápticos. Yang et al. (2014, 12919-20) encontraron una “red sináptica bien organizada, que se proyecta longitudinalmente entre las neuronas piramidales CA1. [y] que las sinapsis de esta red son capaces de soportar la plasticidad sináptica, incluida la potenciación a largo plazo, y un mecanismo de memoria a corto plazo llamado 'retención dendrítica y lectura. '. [Además,] LTP puede fortalecer las conexiones interlamelares de Ca1-a-CA1 así como las conexiones bien establecidas de Ca3 a CA1 en el plano transversal” (Yang et al., 2014, p. 12921). ¿Podrían estas dos redes en planos perpendiculares entre sí en el hipocampo sugerir una especie de disposición en forma de cuadrícula que podría codificar la ubicación de objetos, incluido uno mismo, en el espacio tridimensional? Como se mencionó anteriormente, CA3 está implicado en la representación del espacio y las relaciones espaciales. Además, los neurocientíficos han encontrado “células de lugar” en el hipocampo que se disparan cuando un animal se encuentra en un lugar específico de un laberinto, por ejemplo, y las “células de lugar” sintonizadas para disparar a lugares específicos también parecen estar presentes en el hipocampo humano. Según Yang et al. (2014, p. 12923), “Estas propiedades sugieren que este sistema puede ser un componente integral de la red de procesamiento de información 3D más grande del hipocampo”. Claramente, el procesamiento del espacio tridimensional y las relaciones espaciales de uno con otros objetos es fundamental para navegar por el mundo con éxito.
Sin embargo, Yang y sus colegas consideran otra posibilidad: que la red CA1 pueda “transformar una secuencia de tiempo en una secuencia espacial” de modo que la “transformada resultante de “tiempo a espacio” sea efectivamente un mecanismo de memoria de secuencia eficiente, tal como sería necesario cuando “la información normalmente llega como un tiempo secuencia sobre un intervalo finito y su significado sólo puede revelarse si cada secuencia es vista como un todo y en el orden adecuado. [Además,] estudios conductuales han sugerido que el área CA1 posee la capacidad de memoria secuencial” (Yang et al., 2014, p. 12923). Para entender lo que esto significa, piense en los eventos en el mundo: ocurren a lo largo del tiempo y su significado, incluso a menudo su significado adaptativo, solo puede hacerse evidente cuando se los percibe como una secuencia completa. Si introspectas por un momento y consideras cómo es un recuerdo cuando lo recuerdas, es posible que tengas una idea de esto. Por ejemplo, tengo un recuerdo muy distinto de la primera vez que vi el planeta Saturno y sus anillos a través de un telescopio que compré cuando tenía 14 años (que todavía tengo) usando dinero que había ahorrado al entregar periódicos puerta a puerta en mi ruta de papel (sí, soy tan viejo). Después de lo que ahora es hace 61 años, todavía recuerdo haber instalado mi telescopio en una noche fría en la entrada de la casa de mis padres, encontrar a Saturno casi por accidente, ver los anillos alrededor del planeta y luego correr dentro por la puerta de la cocina para contarles a mis padres lo que estaba viendo. ¿Podría ser que la subregión CA1 del hipocampo sea un componente importante en la capacidad de recordar eventos en su secuencia apropiada? La memoria secuencial parece ser una propiedad esencial del cerebro para representar y almacenar información vital sobre el orden temporal de los eventos que experimentamos en el mundo.
Figura\(\PageIndex{8}\): Diagrama que muestra la anatomía del hipocampo. Las neuronas piramidales en el área CA1 tienen hasta 30,000 espinas dendríticas por neurona en sus dendritas. Las espinas están asociadas con sinapsis que son principalmente excitadoras. La subregión CA3-CA4 del hipocampo se indica en áreas negras, punteadas y eclosionadas. Áreas negras: campos terminales de fibra musgosa suprapiramidal (SP), intra e infrapiramidal (IIP) e hilar (CA4) originados en el giro dentado. Área punteada: estratos oriens (OR) y radiatum (RD). Área eclosionada: estrato lacunosum-moleculare (LM). CA1, subregión del hipocampo sin fibras musgosas; FI, hipocampo fimbria; FD, fascia dentata; OL y ML, capas moleculares externas y medias de la fascia dentata; SG, capa supragranular; GC, células granulares (Imagen y subtítulo de Wikimedia Commons; Archivo:Diagrama de una sección transversal teñida con TIMM del hippocampus.JPEG; https://commons.wikimedia.org/wiki/F...ppocampus.JPEG; por Sluyter, Frans; Laure Jamot, Jean-Yves Bertholet, Wim Crusio (2005-04-22). “La exposición prenatal al alcohol no afecta el aprendizaje del laberinto radial y los tamaños de fibra musgosa del hipocampo en tres cepas endogámicas de ratón”. Funciones Conductuales y Cerebrales 1 (1): 5. DOI: 10.1186/1744-9081-1-5. ISSN 1744-9081. Obtenido el 21-12-2007; licenciado bajo la licencia Creative Commons Attribution 2.0 Generic).
Referencias
Bennett E.L., et al. (1964). Plasticidad química y anatómica del cerebro. Ciencia, 146:610. [PubMed]
Bertling, E., Ludwig, A., Koskinen, M., & Hotulainen, P. (2012). Métodos para el análisis tridimensional de la dinámica de la columna dendrítica. En Métodos en enzimología (Vol. 506, pp. 391-406). Prensa Académica.
Bliss T.V. & Collingridge G.L. (1993). Un modelo sináptico de memoria: potenciación a largo plazo en el hipocampo. Naturaleza. 361 (6407): 31—39
Bliss, T. V., Collingridge, G. L., Morris, R. G., & Reymann, K. G. (2018). Potenciación a largo plazo en el hipocampo: descubrimiento, mecanismos y función. Neuroforo, 24 (3), A103-A120.
Cherubini, E., & Miles, R. M. (2015). La región CA3 del hipocampo: ¿cómo es? ¿Para qué sirve? ¿Cómo lo hace?. Fronteras en la neurociencia celular, 9, 19.
Citri A., & Malenka R.C. (2008). Plasticidad sináptica: múltiples formas, funciones y mecanismos. Neuropsicofarmacología. 33 (1): 18—41.
Colley P.A., Routtenberg A. (1993). Potenciación a largo plazo como diálogo sináptico. Brain Res Brain Res Rev. 18:115 [PubMed]
DeZazzo J., Tully T. (1995). Disección de la formación de memoria: de la farmacología conductual a la genética molecular. Tendencias Neurosci. 18:212. [PubMed]
Hebb, D.O. (1949). La Organización de la Conducta. Nueva York. Wiley.
Irie, F., & Yamaguchi, Y. (2009). Señalización del receptor Eph y morfología de la columna. Enciclopedia de Neurociencia, p. 1141-1145.
Koskinen, M., Bertling, E., & Hotulainen, P. (2012). Métodos para medir la tasa de avance de actina en espinas dendríticas. En Métodos en enzimología (Vol. 505, pp. 47-58). Prensa Académica.
Krasne F.B., Glanzman D.L. (1995). Lo que podemos aprender del aprendizaje de invertebrados. Annu Rev Psychol. 46:585.
Leuner, B., & Shors, T. J. (2010). Formación de sinapsis y memoria. En Enciclopedia de Neurociencia del Comportamiento (pp. 349-355). Elsevier
Mizumori S.J., Rosenzweig M.R., Bennett E.L. (1985). Memoria de trabajo a largo plazo en la rata: efectos de la anisomicina aplicada hipocampalmente. Behav Neurosci.; 99:220. [PubMed]
Nicoll, R. A., & Kauer, J. A. Malenka. RC 1988. La excitación actual en la potenciación a largo plazo. Neurona, 1, 97-103.
Park, P., Georgiou, J., Sanderson, T.M. et al. (2021). PKA impulsa un aumento en la conductancia unitaria del receptor AMPA durante la LTP en el hipocampo. Nat Commun 12, 413. Este artículo está bajo una licencia Creative Commons Attribution 4.0.
Rose S.P.R. (1992). Neuropsicología de la memoria. Escudero L.R., et al., editores. Guilford; Nueva York: p. 547.
Rose S.P. (1995). Glicoproteínas y formación de memoria. Behav Cerebro Res. 66:73. [PubMed]
Rosenzweig M.R., et al. (1992). Neuropsicología de la memoria. Escudero LR, et al., editores. Guilford; Nueva York: p. 533.
Rosenzweig, M. R. (2007). Modificación de circuitos cerebrales a través de la experiencia. En Plasticidad Neural y Memoria: De los Genes a la Imagen Cerebral. CRC Press/Taylor & Francis, Boca Raton (FL); 2007. PMID: 21204433.
Serrano P.A., et al. (1994). Efectos diferenciales de inhibidores y activadores de proteína quinasa sobre la formación de memoria en el pollo de 2 días de edad. Behav Neural Biol. 61:60. [PubMed]
Traynelis S.F., Wollmuth L.P., McBain C.J., Menniti F.S., Vance K.M., Ogden K.K., Hansen K.B., Yuan H., Myers S.J., Dingledine, R. (2010). Canales iónicos receptores de glutamato: estructura, regulación y función. Pharmacol. Rev. 62 (3): 405—496
Tully T, et al. Un retorno a la disección genética de la memoria en Drosophila. Harb de Primavera Fría Symp Quant Biol. 1996; 61:207. [PubMed]
Yang, S., Yang, S., Moreira, T., Hoffman, G., Carlson, G. C., Bender, K. J.,... & Tang, C. M. (2014). Red interlamelar CA1 en el hipocampo. Actas de la Academia Nacional de Ciencias, 111 (35), 12919-12924.
Referencias adicionales
Álvarez, V. A., & Sabatini, B. L. (2007). Plasticidad anatómica y fisiológica de espinas dendríticas. Annú. Rev. Neurosci. , 30, 79-97.
Bliss, T. V., & Lømo, T. (1973). Potenciación de larga duración de la transmisión sináptica en el área dentada del conejo anestesiado tras la estimulación de la trayectoria perforante. La Revista de fisiología, 232 (2), 331-356.
Chidambaram, S. B., Rathipriya, A. G., Bolla, S. R., Bhat, A., Ray, B., Mahalakshmi, A. M.,... & Sakharkar, M. K. (2019). Espinas dendríticas: revisitando el papel fisiológico. Avances en Neuropsicofarmacología y Psiquiatría Biológica, 92, 161-193.
Eichenbaum, H. (1999). La conciencia consciente, la memoria y el hipocampo. Neurociencia de la naturaleza, 2 (9), 775—776.
Hayashi-Takagi, A., Yagishita, S., Nakamura, M., Shirai, F., Wu, Y.I., Loshbaugh, A.L., Kuhlman, B., Hahn, KM., Kasai, H. (2015). Etiquetado y borrado óptico de trazas de memoria sináptica en la corteza motora. Naturaleza 525, 333—338.
Harris, K. M., Fiala, J. C., & Ostroff, L. (2003). Cambios estructurales en las sinapsis de columna dendrítica durante la potenciación a largo plazo. Transacciones filosóficas de la Royal Society de Londres. Serie B: Ciencias Biológicas, 358 (1432), 745-748.
Hebb, D.O. (1949). La Organización de la Conducta. Nueva York. Wiley.
Henry, F. E., Hockeimer, W., Chen, A., Mysore, S. P., & Sutton, M. A. (2017). La diana mecanicista de la rapamicina es necesaria para cambios en la morfología de la columna dendrítica asociados con la potenciación a largo Cerebro Molecular, 10 (1), 1-17.
Jonides, J., Lacey, S. C., & Nee, D. E. (2005). Procesos de memoria de trabajo en mente y cerebro. Direcciones Actuales en Ciencia Psicológica, 14 (1), 2—5.
Kaada, B. R., Rasmussen, E. W., & Kveim, O. (1961). Efectos de las lesiones hipocampales sobre el aprendizaje y retención de laberinto en ratas. Neurología Experimental, 3 (4), 333-355.
Kandel, E. (1976). Bases de Comportamiento Celular. San Francisco. W.H. Freeman y Compañía.
Kandel, E. R., & Schwartz, J. H. (1982). Biología molecular del aprendizaje: Modulación de la liberación del transmisor. Ciencia, 218 (4571), 433—443
Kasai, Haruo; Matsuzaki, Masanori; Noguchi, Jun; Yasumatsu, Nobuaki; Nakahara, Hiroyuki (1° de julio de 2003). “Relaciones estructura-estabilidad—función de espinas dendríticas”. Tendencias en Neurociencias. 26 (7): 360—368.
Kasthuri, N., Hayworth, K. J., Berger, D. R., Schalek, R. L., Conchello, J. A., Knowles-Barley, S.,... & Lichtman, J. W. (2015). Reconstrucción saturada de un volumen de neocórtex. Celda, 162 (3), 648-661.
Kim, C. H., & Lisman, J. E. (1999). Un papel del filamento de actina en la transmisión sináptica y la potenciación a largo plazo. Revista de Neurociencia, 19 (11), 4314-4324.
Kolb, B. & Whishaw, I.Q. (2001). En Introducción al Cerebro y Comportamiento. Nueva York. Worth Publishers.
Krucker, T., Siggins, G. R., & Halpain, S. (2000). Los filamentos dinámicos de actina son necesarios para la potenciación estable a largo plazo (LTP) en el área CA1 del hipocampo. Actas de la Academia Nacional de Ciencias, 97 (12), 6856-6861.
Krupa, D. J., Thompson, J. K., & Thompson, R. F. (1993). Localización de un rastro de memoria en el cerebro de mamíferos. Ciencia, 260 (5110), 989—991.
Lashley, K. S. (1929). Los efectos de las lesiones cerebrales posteriores a la formación del hábito de laberinto: Localización del hábito. En Mecanismos cerebrales e inteligencia: Un estudio cuantitativo de las lesiones cerebrales (pp. 86—108). Chicago, IL: Prensa de la Universidad de Chicago.
Miller, R. R., & Marlin, N. A. (2014). Amnesia tras choque electroconvulsivo. Trastornos funcionales de la memoria, 143-178. Nueva York. Prensa de Psicología.
Murakoshi, H., Wang, H., & Yasuda, R. (2011). Activación local y persistente de GTPasas Rho durante la plasticidad de espinas dendríticas individuales. Naturaleza, 472 (7341), 100-104.
Ofer, N., Berger, D. R., Kasthuri, N., Lichtman, J. W., & Yuste, R. (2021). El análisis ultraestructural de cuellos de columna dendrítica revela un continuo de morfologías espinales. Neurobiología del Desarrollo.
Pinel, P.J. & Barnes, S. (2021). Biopsicología (11ª Edición). Boston. Educación Pearson.
Sigurdsson, T., Doyère, V., Caín, C. K., & LeDoux, J. E. (2007). Potenciación a largo plazo en la amígdala: Un mecanismo celular de aprendizaje y memoria por miedo. Neurofarmacología, 52 (1), 215—227.
Escudero, L. R. (1992). La memoria y el hipocampo: Una síntesis a partir de hallazgos con ratas, monos y humanos. Revisión Psicológica, 99 (2), 195—231.
Thompson, R. (1974). Localización del “sistema de memoria laberinto” en la rata blanca. Psicología Fisiológica, 2 (1), 1-17.
Woodruff-Pak, D. S., Goldenberg, G., Downey-Lamb, M. M., Boyko, O. B., & Lemieux, S. K. (2000). Volumen cerebeloso en humanos relacionado con la magnitud del condicionamiento clásico. Neuroreport: Para la comunicación rápida de la investigación en neurociencias, 11 (3), 609—615.
Yang, G., Pan, F., & Gan, W. B. (2009). Las espinas dendríticas mantenidas de manera estable están asociadas con recuerdos de por vida. Naturaleza, 462 (7275), 920-924.
Atribuciones
“Objetivos de aprendizaje”, “Visión general”, “La búsqueda del aprendizaje y la memoria en la sinapsis”, “Cambios en la sinapsis correlacionados con el aprendizaje y la memoria”, “Cambios en la sinapsis correlacionados con el aprendizaje y la memoria: cambios anatómicos en las espinas dendríticas”, “LTP, cascadas neuroquímicas y etapas de la memoria”, original material escrito por Kenneth A. Koenigshofer, Ph.D., está licenciado bajo CC BY 4.0.
“Plasticidad sináptica”, “Plasticidad sináptica: potenciación a corto plazo, potenciación a largo plazo y depresión a largo plazo”, “Mejora sináptica a corto plazo y depresión”, “Potenciación a largo plazo (LTP)” y “Depresión a largo plazo (LTD)”, adaptada por Kenneth A. Koenigshofer, Ph.D., de Lumen Boundless Biology, How Neuronas Comunicar; https://courses.lumenlearning.com/bo...s-communicate/; curación y revisión. Proporcionado por: Boundless.com. Licencia: CC BY-SA: Atribución-CompartirIgual.
“Mecanismos de Nivel Celular y Receptor Revisitados” adaptado de El Cerebro de Arriba a Abajo; licencia: Copyleft, https://thebrain.mcgill.ca/flash/pop... /pop_copy.html; modificado por Kenneth A. Koenigshofer, PhD., licenciado bajo CC BY 4.0.