
15.3: Ciclos de Nutrimentos
- Page ID
- 58508

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Explicar los procesos de ciclo de carbono, nitrógeno y fósforo.
- Explicar el proceso por el cual los rizobios infectan leguminosas y forman nódulos radiculares.
Los ciclos de nutrientes, también conocidos como ciclos biogeoquímicos, describen el movimiento de elementos químicos a través de diferentes medios, como la atmósfera, el suelo, las rocas, los cuerpos de agua y los organismos. Los ciclos de nutrientes mantienen los elementos esenciales disponibles para las plantas y otros organismos. A continuación se discute el ciclo de tres macronutrientes. El primero es el ciclo del carbono. Las plantas adquieren carbono a través de la fotosíntesis. También se discuten los ciclos de nitrógeno y fósforo, y las plantas adquieren nitrógeno y fósforo como nutrientes minerales del suelo.
El ciclo del carbono
El ciclo del carbono en realidad está compuesto por varios ciclos interconectados: uno que se ocupa del intercambio rápido de carbono entre organismos vivos y el otro con el ciclo a largo plazo del carbono a través de procesos geológicos (Figura\(\PageIndex{1}\)). El efecto general es que el carbono se recicla constantemente en los procesos dinámicos que tienen lugar en la atmósfera, en la superficie y en la corteza terrestre. La gran mayoría del carbono reside como minerales inorgánicos en las rocas de la corteza. Otros reservorios de carbono, lugares donde se acumula el carbono, incluyen los océanos y la atmósfera. Algunos de los átomos de carbono en su cuerpo hoy en día pueden haber residido hace mucho tiempo en el cuerpo de un dinosaurio, o quizás alguna vez fueron enterrados en lo profundo de la corteza terrestre como minerales de roca carbonatada.
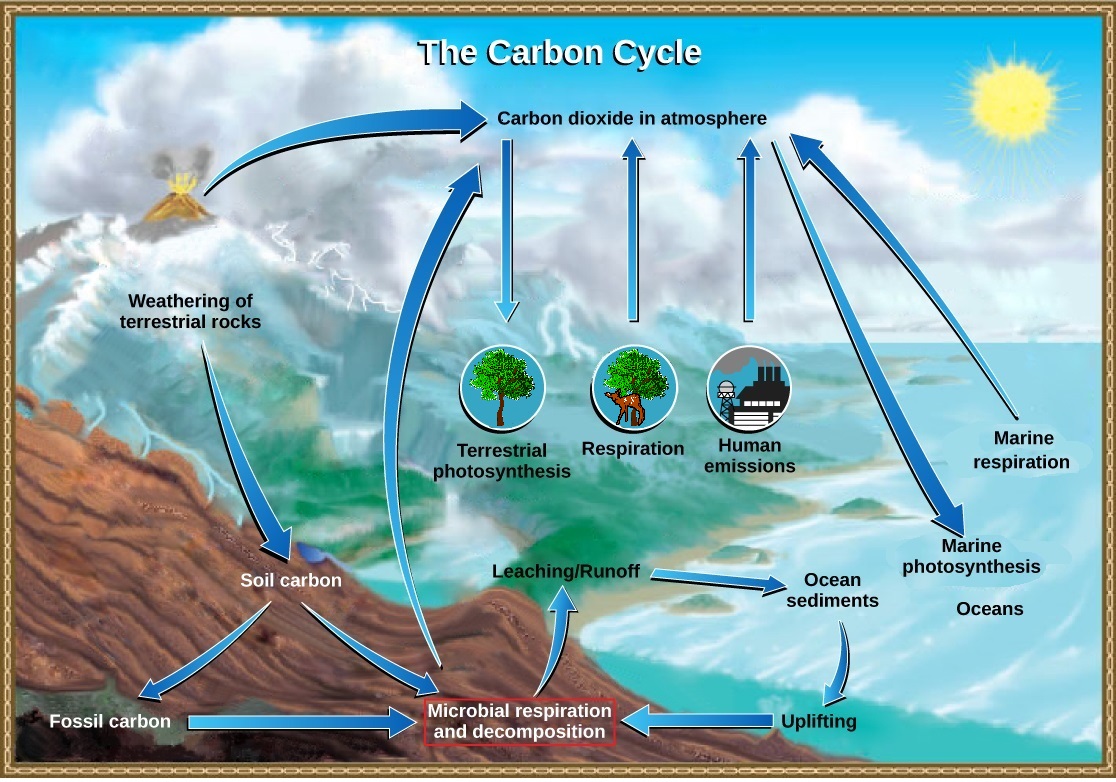
El carbono circula lentamente entre la tierra y el océano
En tierra, el carbono se almacena en el suelo como carbono orgánico en forma de organismos en descomposición o rocas terrestres. Las plantas y algas descompuestas a veces son enterradas y comprimidas entre capas de sedimentos. Después de millones de años se forman combustibles fósiles como el carbón, el petróleo y el gas natural. La meteorización de rocas terrestres y minerales liberan carbono en el suelo.
Los compuestos que contienen carbono en el suelo se pueden lavar en cuerpos de agua a través de la lixiviación. Esta agua finalmente ingresa al océano. El dióxido de carbono atmosférico también se disuelve en el océano, reaccionando con moléculas de agua para formar iones carbonato (CO 3 2-). Algunos de estos iones se combinan con iones de calcio en el agua de mar para formar carbonato de calcio (CaCo 3), un componente importante de las conchas de los organismos marinos. Estos organismos finalmente mueren y sus conchas forman sedimentos en el fondo oceánico. A lo largo del tiempo geológico, el carbonato de calcio forma piedra caliza, que comprende el mayor reservorio de carbono de la Tierra
El carbonato también precipita en sedimentos, formando rocas carbonatadas, como la piedra caliza. Los sedimentos de carbono del fondo oceánico son capturados profundamente dentro de la Tierra por el proceso de subducción: el movimiento de una placa tectónica debajo de otra. Los sedimentos oceánicos son subducidos por las acciones de la tectónica de placas, fundidos y luego regresados a la superficie durante la actividad volcánica. La tectónica de placas también puede causar edificante y devolver los sedimentos oceánicos a la tierra.
Ciclos de carbono rápidamente entre los organismos y la atmósfera
El dióxido de carbono se convierte en glucosa, una molécula orgánica rica en energía a través de la fotosíntesis de plantas, algas y algunas bacterias. Luego pueden producir otras moléculas orgánicas como carbohidratos complejos (como el almidón), proteínas y lípidos, que los animales pueden comer. La mayoría de los autótrofos terrestres obtienen su dióxido de carbono directamente de la atmósfera, mientras que los autótrofos marinos lo adquieren en forma disuelta (bicarbonato, HCO 3 —).
Plantas, animales y otros organismos descomponen estas moléculas orgánicas durante el proceso de respiración celular aeróbica, que consume oxígeno y libera energía, agua y dióxido de carbono. El dióxido de carbono se devuelve a la atmósfera durante el intercambio gaseoso. Otro proceso por el que se recicla el material orgánico es la descomposición de organismos muertos. Durante este proceso, bacterias y hongos descomponen los compuestos orgánicos complejos. Los descomponedores pueden hacer respiración, liberar dióxido de carbono u otros procesos que liberen metano (CH 4). La fotosíntesis y la respiración son en realidad recíprocas entre sí con respecto al ciclo del carbono: la fotosíntesis elimina el dióxido de carbono de la atmósfera y la respiración lo devuelve. Por lo tanto, una interrupción significativa de un proceso puede afectar la cantidad de dióxido de carbono en la atmósfera.
La respiración celular es solo un proceso que libera dióxido de carbono. Los procesos físicos, como la erupción de volcanes y la liberación de respiraderos hidrotermales (aberturas en el fondo oceánico) agregan dióxido de carbono a la atmósfera. Adicionalmente, la combustión de madera y combustibles fósiles libera dióxido de carbono. El nivel de dióxido de carbono en la atmósfera está muy influenciado por el reservorio de carbono en los océanos. El intercambio de carbono entre la atmósfera y los reservorios de agua influye en la cantidad de carbono que se encuentra en cada uno.
Ciclo de Nitrógeno
Es difícil introducir nitrógeno en los organismos vivos. Las plantas y las algas no están equipadas para incorporar nitrógeno de la atmósfera (donde existe como N 2 covalente fuertemente unido, triple covalente) aunque esta molécula comprende aproximadamente el 78 por ciento de la atmósfera. Debido a que la mayor parte del nitrógeno se almacena en la atmósfera, la atmósfera se considera un reservorio de nitrógeno.
Fijación de nitrógeno
La molécula de nitrógeno (N 2) es bastante inerte. Para descomponerlo para que sus átomos puedan combinarse con otros átomos requiere la entrada de cantidades sustanciales de energía. La fijación de nitrógeno es el proceso de convertir gas nitrógeno en amoníaco (NH 3), que espontáneamente se convierte en amonio (NH 4 +). El amonio se encuentra en cuerpos de agua y en el suelo (Figura\(\PageIndex{2}\)).

Tres procesos son responsables de la mayor parte de la fijación de nitrógeno en la biosfera. El primero es la fijación atmosférica por un rayo. La enorme energía de los rayos rompe las moléculas de nitrógeno y permite que sus átomos se combinen con el oxígeno del aire formando óxidos de nitrógeno. Estos se disuelven en la lluvia, formando nitratos, que son llevados a la tierra. La fijación de nitrógeno atmosférico probablemente aporta alrededor del 5-8% del nitrógeno total fijado. El segundo proceso es la fijación industrial. Bajo gran presión, a una temperatura de 600°C, y con el uso de un catalizador, se pueden combinar nitrógeno atmosférico e hidrógeno (generalmente derivado de gas natural o petróleo) para formar amoníaco (NH 3). El amoníaco se puede utilizar directamente como fertilizante, pero la mayor parte se procesa posteriormente a urea y nitrato de amonio (NH 4 NO 3).
El tercer proceso es la fijación biológica. Algunas bacterias fijadoras de nitrógeno, como Azotobacter, son de vida libre. Otros (Rhizobium y Bradyrhizobium) forman una relación simbiótica con las plantas de Fabaceae (familia de frijol o leguminosas), que incluye frijol, guisantes, soja, alfalfa, tréboles y muchas otras especies (Figura\(\PageIndex{3}\)). Las bacterias que forman nódulos radiculares en las leguminosas se denominan informalmente rizobios. Frankia forma nódulos radiculares en alisos, que son árboles no leguminosos. Las cianobacterias fijadoras de nitrógeno que son simbióticas con el helecho acuático Azolla son esenciales para mantener la fertilidad de ambientes semiacuáticos como arrozales. Los líquenes que contienen cianobacterias también pueden fijar el nitrógeno. La fijación biológica de nitrógeno requiere un complejo conjunto de enzimas y un enorme gasto de ATP. Aunque el primer producto estable del proceso es el amoníaco, éste se incorpora rápidamente en proteínas y otros compuestos orgánicos nitrogenados.

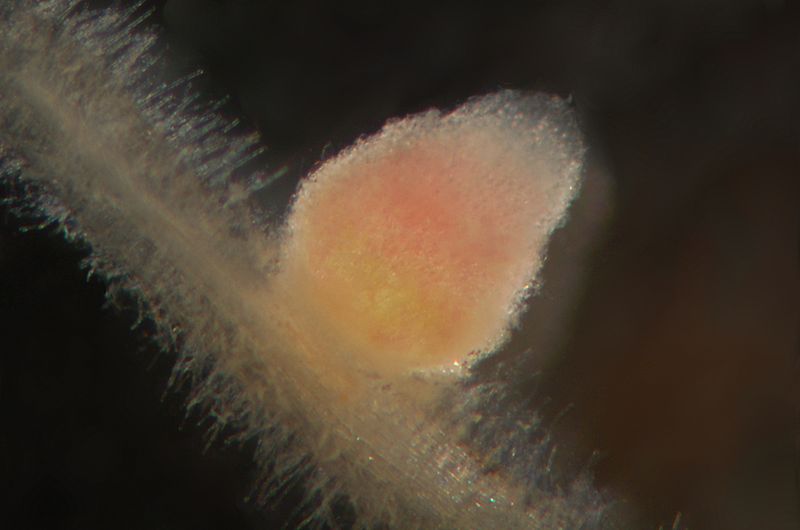
Los rizobios viven libremente en el suelo, pero no pueden fijar el nitrógeno atmosférico hasta que hayan infectado las raíces de una leguminosa. Las legumbres aportan carbohidratos a las bacterias fijadoras de nitrógeno para obtener energía y limitan el oxígeno en el nódulo radicular. La nitrogenasa, la enzima que fija el nitrógeno, es inhibida por el oxígeno, pero los bacteroides aún necesitan algo de oxígeno para conducir la respiración celular y producir ATP. Las legumbres producen leghemoglobina, que se une al oxígeno como la hemoglobina de los vertebrados. Esto reduce la disponibilidad de oxígeno en el nódulo radicular. La leghemoglobina contiene hierro y aparece roja cuando se une al oxígeno; de hecho, un nódulo recién cortado es rojo o rosado (Figura\(\PageIndex{4}\)).
Los pelos radiculares liberan químicos llamados flavonoides, que hacen que las bacterias sinteticen factores Nod. Estas son señales químicas que se unen a los receptores de células epidérmicas radiculares e inducen a la leguminosa a producir nódulos. Los factores de cabeceo median las interacciones entre cepas específicas de rizobios y las especies leguminosas asociadas. Diferentes cepas de rizobios producen diferentes factores Nod, y diferentes leguminosas producen receptores de diferente especificidad. Debido a la especificidad de la interacción entre el factor Nod y el receptor sobre la leguminosa, algunas cepas de rizobios infectarán solo a guisantes, algunas solo trébol, algunas solo alfalfa, etc. El tratamiento de semillas de leguminosas con la cepa adecuada de rizobia es una práctica agrícola rutinaria.
Si la combinación es correcta, las bacterias ingresan a una célula epidérmica de la raíz y luego migran a la corteza. Su trayectoria discurre dentro de un canal intracelular que crece a través de una célula de la corteza tras otra. Este hilo de infección está construido por el pelo de la raíz, no por las bacterias, y se forma solo en respuesta a la infección. Cuando el hilo de infección alcanza una célula profunda en la corteza, estalla y los rizobios son engullidos por la endocitosis en simbiosomas encerrados en la membrana dentro del citoplasma. En este momento la célula cortical (corteza) pasa por varias rondas de mitosis, sin citocinesis, por lo que la célula se vuelve poliploide. Luego, las células corticales comienzan a dividirse rápidamente formando un nódulo. Esta respuesta es impulsada por la translocación de citoquininas de las células epidérmicas a las células de la corteza. Los rizobios también pasan por un período de rápida multiplicación dentro de las células del nódulo. Entonces empiezan a cambiar de forma y pierden su motilidad. Los bacteroides, como ahora se les llama, casi pueden llenar la célula. Sólo ahora comienza la fijación de nitrógeno.
Nitrificación
El amonio es convertido por bacterias y arqueas en nitritos (NO 2 −) y luego nitratos (NO 3 −) a través del proceso de nitrificación. Al igual que el amonio, los nitritos y nitratos se encuentran en el agua y el suelo.
Desnitrificación
Algunos nitratos se convierten de nuevo en gas nitrógeno, que se libera a la atmósfera. El proceso, llamado desnitrificación, es conducido por bacterias.
Asimilación
El amonio y los nitratos pueden ser utilizados directamente por las plantas y otros productores para elaborar moléculas orgánicas como el ADN y las proteínas a través del proceso de asimilación. Este nitrógeno ya está disponible para los consumidores. El nitrógeno orgánico es especialmente importante para el estudio de la dinámica de los ecosistemas porque muchos procesos, como la producción primaria, están limitados por el suministro disponible de nitrógeno.
Amonificación
Los consumidores excretan compuestos orgánicos nitrogenados que regresan al ambiente. Adicionalmente, los organismos muertos en cada nivel trófico contienen nitrógeno orgánico. Los microorganismos, como bacterias y hongos, descomponen estos desechos y tejidos muertos, produciendo finalmente amonio a través del proceso de amonificación.
Ciclo Fósforo
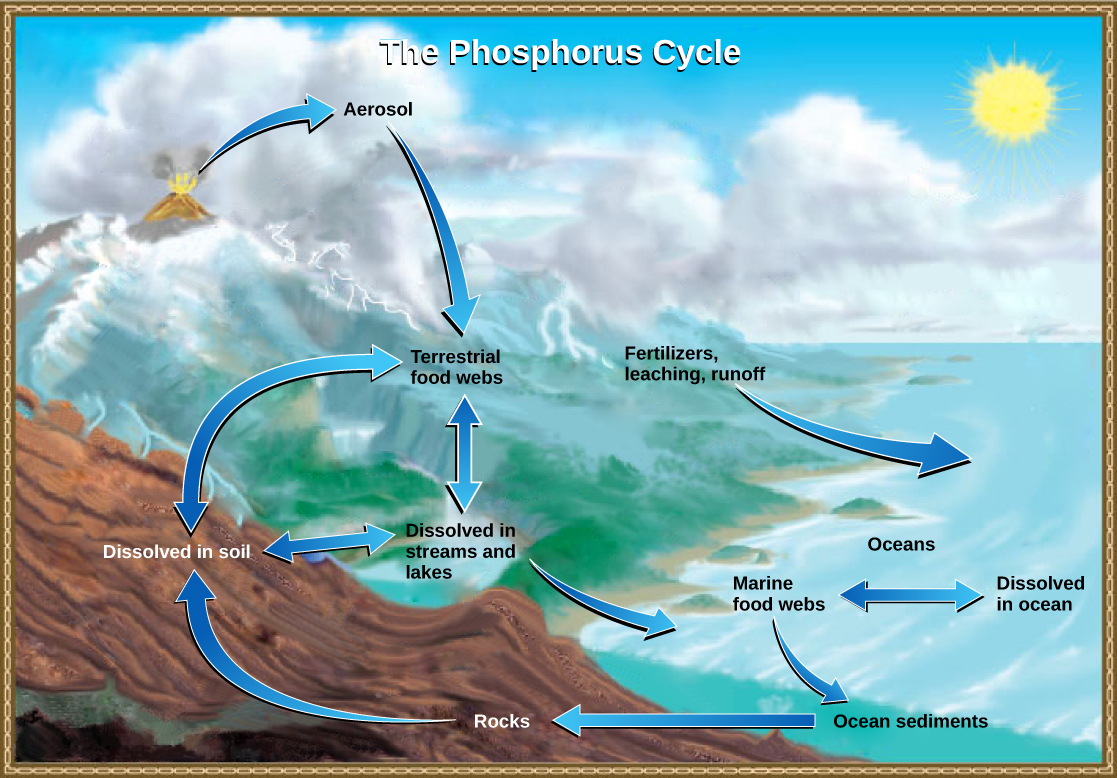
Varias formas de nitrógeno (N 2, NH 4 +, NO 3 −, etc.) estuvieron involucradas en el ciclo del nitrógeno, pero el fósforo permanece principalmente en forma del ion fosfato (PO 4 3-). También en contraste con el ciclo del nitrógeno, no hay forma de fósforo en la atmósfera.
Las rocas son un reservorio de fósforo, y estas rocas tienen su origen en el océano. Los sedimentos oceánicos que contienen fosfato se forman principalmente a partir de los cuerpos de organismos marinos y de sus excreciones. Sin embargo, la ceniza volcánica, los aerosoles y el polvo mineral también pueden ser fuentes significativas de fosfato. Este sedimento se mueve luego a la tierra a lo largo del tiempo geológico por la elevación de la superficie de la Tierra (Figura\(\PageIndex{5}\)). El movimiento del fosfato del océano a la tierra y a través del suelo es extremadamente lento, teniendo el ion fosfato promedio un tiempo de residencia oceánica entre 20,000 y 100,000 años.
La meteorización de las rocas libera fosfatos en el suelo y cuerpos de agua. Las plantas pueden asimilar fosfatos en el suelo e incorporarlos en moléculas orgánicas, poniendo el fósforo a disposición de los consumidores en las redes alimentarias terrestres. Los desechos y los organismos muertos son descompuestos por hongos y bacterias, liberando fosfatos de nuevo al suelo. Algo de fosfato se lixivia del suelo, entrando en ríos, lagos y el océano. Los productores primarios en redes alimentarias acuáticas, como algas y bacterias fotosintéticas, asimilan fosfato, y el fosfato orgánico está así disponible para los consumidores en redes alimentarias acuáticas. Similar a las redes alimentarias terrestres, el fósforo se intercambia recíprocamente entre el fosfato disuelto en el océano y el fósforo orgánico en organismos marinos.
Atribuciones
Comisariada y autoría de Melissa Ha utilizando las siguientes fuentes:
- 3.2 Ciclos biogeoquímicos, 2.2 energía y 2.4 energía ingresan a los ecosistemas a través de la fotosíntesis desde la biología ambiental por Matthew R. Fisher (licenciado bajo CC-BY)
- Ciclo del Carbono y Ciclo del Nitrógeno de Biología por John W. Kimball (licenciado bajo CC-BY)
- 6.2 Ciclos biogeoquímicos y el flujo de energía en el sistema terrestre desde la sustentabilidad: una fundación integral por Tom Theis y Jonathan Tomkin, editores (licenciados bajo CC-BY). Descárgala gratis en CNX.
- 2 Cycling of Matter de AP Environmental Science por University of California College Prep, University of California (licenciado bajo CC-BY). Descárgala gratis en CNX.