
17.2: Defensas Químicas
- Page ID
- 54365

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Describir cómo las enzimas en los fluidos corporales brindan protección contra infecciones o enfermedades
- Enumerar y describir la función de péptidos antimicrobianos, componentes del complemento, citocinas y proteínas de fase aguda
- Describir similitudes y diferencias entre las vías clásicas, alternas y de complemento de lectinas
Además de las defensas físicas, el sistema inmune innato inespecífico utiliza una serie de mediadores químicos que inhiben a los invasores microbianos. El término “mediadores químicos” abarca una amplia gama de sustancias que se encuentran en diversos fluidos corporales y tejidos en todo el cuerpo. Los mediadores químicos pueden trabajar solos o conjuntamente entre sí para inhibir la colonización microbiana y la infección.
Algunos mediadores químicos son producidos endógenamente, es decir, son producidos por células del cuerpo humano; otros son producidos exógenamente, es decir, que son producidos por ciertos microbios que forman parte del microbioma. Algunos mediadores se producen continuamente, bañando la zona en la sustancia antimicrobiana; otros se producen o activan principalmente en respuesta a algún estímulo, como la presencia de microbios.
Mediadores químicos y enzimáticos encontrados en fluidos corporales
Los fluidos producidos por la piel incluyen ejemplos de mediadores tanto endógenos como exógenos. Las glándulas sebáceas en la dermis secretan un aceite llamado sebo que se libera sobre la superficie de la piel a través de los folículos pilosos. Este sebo es un mediador endógeno, proporcionando una capa adicional de defensa al ayudar a sellar el poro del folículo piloso, evitando que las bacterias en la superficie de la piel invadan las glándulas sudoríparas y el tejido circundante (Figura\(\PageIndex{1}\)). Ciertos miembros del microbioma, como la bacteria Propionibacterium acnes y el hongo Malassezia, entre otros, pueden utilizar enzimas lipasa para degradar el sebo, usándolo como fuente de alimento. Esto produce ácido oleico, que crea un ambiente ligeramente ácido en la superficie de la piel que es inhóspito para muchos microbios patógenos. El ácido oleico es un ejemplo de un mediador producido exógenamente porque es producido por microbios residentes y no directamente por las células del cuerpo.
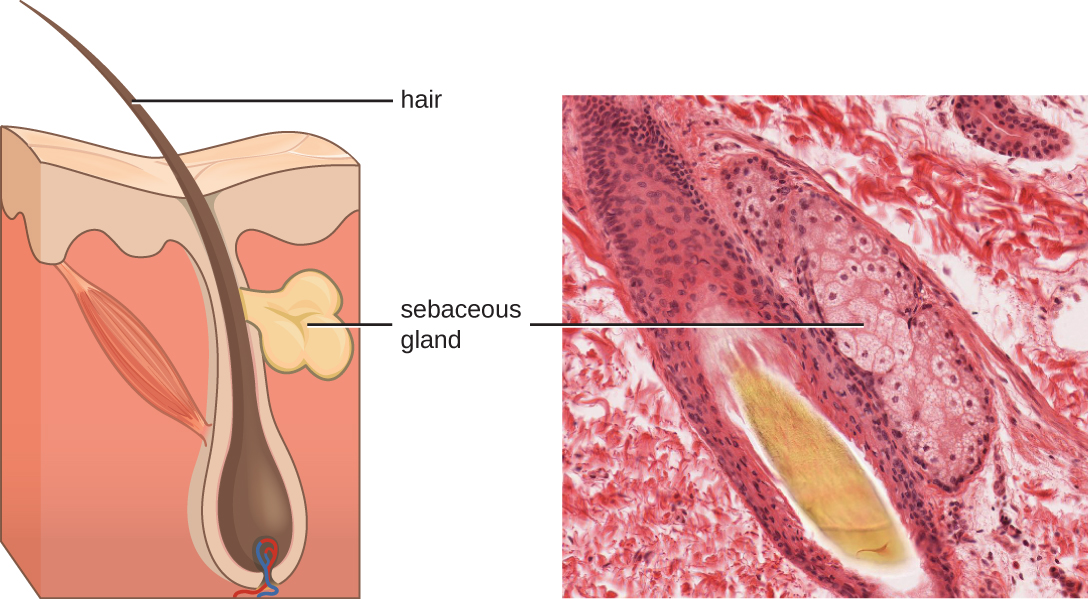
Los factores ambientales que afectan la microbiota de la piel pueden tener un impacto directo en la producción de mediadores químicos. La baja humedad o la disminución de la producción de sebo, por ejemplo, podrían hacer que la piel sea menos habitable para microbios que producen ácido oleico, haciendo así que la piel sea más susceptible a patógenos normalmente inhibidos por el bajo pH de la piel. Muchos humectantes de la piel están formulados para contrarrestar tales efectos restaurando la humedad y los aceites esenciales en la piel.
El tracto digestivo también produce una gran cantidad de mediadores químicos que inhiben o matan microbios. En la cavidad oral, la saliva contiene mediadores como las enzimas lactoperoxidasas, y el moco secretado por el esófago contiene la enzima antibacteriana lisozima. En el estómago, el líquido gástrico altamente ácido mata a la mayoría de los microbios. En el tracto digestivo inferior, los intestinos tienen enzimas pancreáticas e intestinales, péptidos antibacterianos (criptinas), bilis producida a partir del hígado y células Paneth especializadas que producen lisozima. Juntos, estos mediadores son capaces de eliminar la mayoría de los patógenos que logran sobrevivir al ambiente ácido del estómago.
En las vías urinarias, la orina elimina los microbios del cuerpo durante la micción. Además, la ligera acidez de la orina (el pH promedio es de aproximadamente 6) inhibe el crecimiento de muchos microbios y patógenos potenciales en el tracto urinario.
El sistema reproductivo femenino emplea lactato, un mediador químico producido exógenamente, para inhibir el crecimiento microbiano. Las células y capas de tejido que componen la vagina producen glucógeno, un polímero ramificado y más complejo de glucosa. Los lactobacilos de la zona fermentan glucógeno para producir lactato, disminuyendo el pH en la vagina e inhibiendo la microbiota transitoria, patógenos oportunistas como Candida (una levadura asociada a infecciones vaginales) y otros patógenos responsables de enfermedades de transmisión sexual.
En los ojos, las lágrimas contienen los mediadores químicos lisozima y lactoferrina, ambos de los cuales son capaces de eliminar microbios que han encontrado su camino hacia la superficie de los ojos. La lisozima escinde el enlace entre NAG y NAM en peptidoglicano, un componente de la pared celular en bacterias. Es más eficaz contra las bacterias grampositivas, que carecen de la membrana externa protectora asociada a las bacterias gramnegativas. La lactoferrina inhibe el crecimiento microbiano mediante la unión química y el secuestro de hierro. Esto efectivamente muere de hambre a muchos microbios que requieren hierro para su crecimiento.
En los oídos, el cerumen (cerumen) exhibe propiedades antimicrobianas debido a la presencia de ácidos grasos, los cuales bajan el pH entre 3 y 5.
El tracto respiratorio utiliza diversos mediadores químicos en las fosas nasales, tráquea y pulmones. El moco producido en las fosas nasales contiene una mezcla de moléculas antimicrobianas similares a las que se encuentran en las lágrimas y la saliva (por ejemplo, lisozima, lactoferrina, lactoperoxidasa). Las secreciones en la tráquea y los pulmones también contienen lisozima y lactoferrina, así como un grupo diverso de mediadores químicos adicionales, como el complejo lipoproteico llamado surfactante, que tiene propiedades antibacterianas.
Ejercicio\(\PageIndex{1}\)
- Explicar la diferencia entre mediadores endógenos y exógenos
- Describir cómo el pH afecta las defensas antimicrobianas
Péptidos antimicrobianos
Los péptidos antimicrobianos (AMP) son una clase especial de mediadores no específicos derivados de células con propiedades antimicrobianas de amplio espectro. Algunos AMP son producidos rutinariamente por el cuerpo, mientras que otros se producen principalmente (o se producen en mayores cantidades) en respuesta a la presencia de un patógeno invasor. La investigación ha comenzado a explorar cómo se pueden usar los AMP en el diagnóstico y tratamiento de enfermedades.
Los AMP pueden inducir daño celular en microorganismos de diversas maneras, incluyendo infligiendo daño a las membranas, destruyendo ADN y ARN, o interfiriendo con la síntesis de la pared celular. Dependiendo del mecanismo antimicrobiano específico, un AMP particular puede inhibir solo ciertos grupos de microbios (por ejemplo, bacterias grampositivas o gramnegativas) o puede ser más ampliamente eficaz contra bacterias, hongos, protozoos y virus. Muchos AMP se encuentran en la piel, pero también se pueden encontrar en otras regiones del cuerpo.
Una familia de AMP llamadas defensinas puede ser producida por células epiteliales en todo el cuerpo así como por defensas celulares como macrófagos y neutrófilos (ver Defensas celulares). Las defensinas pueden ser secretadas o actuar dentro de las células hospedadoras; combaten los microorganismos dañando sus membranas plasmáticas. Los AMP llamados bacteriocinas son producidos exógenamente por ciertos miembros de la microbiota residente dentro del tracto gastrointestinal. Los genes que codifican estos tipos de AMP a menudo se transportan en plásmidos y se pueden pasar entre diferentes especies dentro de la microbiota residente a través de transferencia génica lateral u horizontal.
Hay muchos otros AMP en todo el cuerpo. Las características de algunas de las AMP más significativas se resumen en la Tabla\(\PageIndex{1}\).
| AMP | Secretado por | Sitio del cuerpo | Patógenos inhibidos | Modo de acción |
|---|---|---|---|---|
| Bacteriocinas | Microbiota residente | Tracto gastrointestinal | Bacterias | Interrumbe la membrana |
| Catelicidina | Células epiteliales, macrófagos y otros tipos de células | Piel | Bacterias y hongos | Interrumpe la membrana |
| Defensinas | Células epiteliales, macrófagos, neutrófilos | En todo el cuerpo | Hongos, bacterias y muchos virus | Interrumbe la membrana |
| Dermicidina | Glándulas sudoríparas | Piel | Bacterias y hongos | Interrumpe la integridad de membrana y los canales iónicos |
| Histatinas | Glándulas salivales | Cavidad oral | Hongos | Disperturar la función |
Ejercicio\(\PageIndex{2}\)
¿Por qué los péptidos antimicrobianos (AMP) se consideran defensas inespecíficas?
Mediadores de Proteína Plasma
Muchos factores inmunes innatos inespecíficos se encuentran en el plasma, la porción líquida de la sangre. El plasma contiene electrolitos, azúcares, lípidos y proteínas, cada uno de los cuales ayuda a mantener la homeostasis (es decir, el funcionamiento interno estable del cuerpo), y contiene las proteínas involucradas en la coagulación de la sangre. Las proteínas adicionales que se encuentran en el plasma sanguíneo, como las proteínas de fase aguda, las proteínas del complemento y las citocinas, están involucradas en la respuesta inmune innata inespecífica.
Plasma versus Suero
Hay dos términos para la porción líquida de la sangre: plasma y suero. ¿En qué se diferencian si ambos son fluidos y carecen de células? La porción líquida de la sangre que queda después de que se ha producido la coagulación (coagulación de las células sanguíneas) es el suero. Aunque moléculas como muchas vitaminas, electrolitos, ciertos azúcares, proteínas del complemento y anticuerpos aún están presentes en el suero, los factores de coagulación se agotan en gran medida. El plasma, a la inversa, todavía contiene todos los elementos coagulantes. Para obtener plasma de la sangre, se debe utilizar un anticoagulante para prevenir la coagulación. Ejemplos de anticoagulantes incluyen heparina y ácido etilendiaminotetraacético (EDTA). Debido a que se inhibe la coagulación, una vez obtenida, la muestra debe girarse suavemente en una centrífuga. Las células sanguíneas más pesadas y densas forman un sedimento en el fondo de un tubo de centrífuga, mientras que la porción de plasma fluido, que es más ligera y menos densa, permanece por encima del sedimento celular.
Proteínas de fase aguda
Las proteínas de fase aguda son otra clase de mediadores antimicrobianos. Las proteínas de fase aguda se producen principalmente en el hígado y se secretan en la sangre en respuesta a moléculas inflamatorias del sistema inmune. Los ejemplos de proteínas de fase aguda incluyen proteína C reactiva, amiloide A sérico, ferritina, transferrina, fibrinógeno y lectina de unión a manosa. Cada una de estas proteínas tiene una estructura química diferente e inhibe o destruye microbios de alguna manera (Tabla\(\PageIndex{1}\)).
| Algunas proteínas de fase aguda y sus funciones | |
|---|---|
| Proteína C reactiva | Recubre bacterias (opsonización), preparándolas para la ingestión por fagocitos |
| Amiloide A sérico | |
| Ferritina | Enlazar y secuestrar hierro, inhibiendo así el crecimiento de patógenos |
| Transferrina | |
| Fibrinógeno | Involucrados en la formación de coágulos sanguíneos que atrapan patógenos bacterianos |
| Lectina de unión a manosa | Activa la cascada del complemento |
El Sistema Complemento
El sistema del complemento es un grupo de mediadores de proteínas plasmáticas que pueden actuar como una defensa innata inespecífica al tiempo que sirven para conectar la inmunidad innata y adaptativa (discutido en el siguiente capítulo). El sistema del complemento está compuesto por más de 30 proteínas (incluyendo C1 a C9) que normalmente circulan como proteínas precursoras en la sangre. Estas proteínas precursoras se activan cuando son estimuladas o desencadenadas por una variedad de factores, incluyendo la presencia de microorganismos. Las proteínas del complemento se consideran parte de la inmunidad innata inespecífica porque siempre están presentes en la sangre y los fluidos tisulares, permitiendo que se activen rápidamente. Además, cuando se activan a través de la vía alternativa (descrita más adelante en esta sección), las proteínas del complemento se dirigen a patógenos de manera inespecífica.
El proceso por el cual los precursores del complemento circulantes se vuelven funcionales se denomina activación del complemento. Este proceso es una cascada que puede ser desencadenada por uno de los tres mecanismos diferentes, conocidos como vías alternativas, clásicas y lectinas.
La vía alternativa se inicia por la activación espontánea de la proteína C3 del complemento. La hidrólisis de C3 produce dos productos, C3a y C3b. Cuando no hay microbios invasores presentes, C3b se degrada muy rápidamente en una reacción de hidrólisis usando el agua en la sangre. Sin embargo, si hay microbios invasores presentes, C3b se adhiere a la superficie de estos microbios. Una vez unido, C3b reclutará otras proteínas del complemento en una cascada (Figura\(\PageIndex{2}\)).
La vía clásica proporciona un mecanismo más eficiente de activación de la cascada del complemento, pero depende de la producción de anticuerpos por las defensas inmunitarias adaptativas específicas. Para iniciar la vía clásica, un anticuerpo específico primero debe unirse al patógeno para formar un complejo anticuerpo-antígeno. Esto activa la primera proteína en la cascada del complemento, el complejo C1. El complejo C1 es un complejo proteico multiparte, y cada componente participa en la activación completa del complejo general. Después del reclutamiento y activación del complejo C1, las proteínas del complemento de la vía clásica restantes son reclutadas y activadas en una secuencia en cascada (Figura\(\PageIndex{2}\)).
La vía de activación de lectinas es similar a la ruta clásica, pero se desencadena por la unión de la lectina de unión a manosa, una proteína de fase aguda, a los carbohidratos en la superficie microbiana. Al igual que otras proteínas de fase aguda, las lectinas son producidas por las células hepáticas y comúnmente están reguladas al alza en respuesta a las señales inflamatorias recibidas por el cuerpo durante una infección (Figura\(\PageIndex{2}\)).
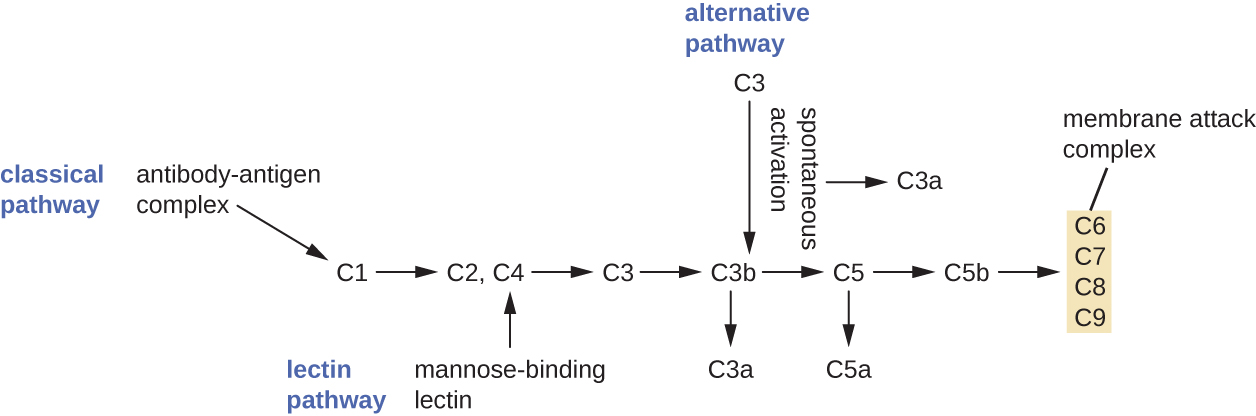
Aunque cada vía de activación del complemento se inicia de una manera diferente, todas proporcionan los mismos resultados protectores: opsonización, inflamación, quimiotaxis y citólisis. El término opsonización se refiere al recubrimiento de un patógeno por una sustancia química (llamada opsonina) que permite a las células fagocíticas reconocerlo, engullirlo y destruirlo más fácilmente. Las opsoninas de la cascada del complemento incluyen C1q, C3b y C4b. Otras opsoninas importantes incluyen proteínas de unión a manosa y anticuerpos. Los fragmentos del complemento C3a y C5a son anafilatoxinas bien caracterizadas con potentes funciones proinflamatorias. Las anafilatoxinas activan los mastocitos, provocando la desgranulación y la liberación de señales químicas inflamatorias, incluyendo mediadores que causan vasodilatación y aumento de la permeabilidad vascular. C5a es también uno de los quimioatrayentes más potentes para neutrófilos y otros glóbulos blancos, defensas celulares que se discutirán en la siguiente sección.
Las proteínas del complemento C6, C7, C8 y C9 se ensamblan en un complejo de ataque a la membrana (MAC), lo que permite que C9 se polimerice en poros en las membranas de bacterias gramnegativas. Estos poros permiten que el agua, los iones y otras moléculas se muevan libremente dentro y fuera de las células diana, lo que eventualmente conduce a la lisis celular y la muerte del patógeno (Figura\(\PageIndex{2}\)). Sin embargo, el MAC solo es efectivo contra bacterias gramnegativas; no puede penetrar la gruesa capa de peptidoglicano asociada a las paredes celulares de bacterias grampositivas. Dado que el MAC no representa una amenaza letal para los patógenos bacterianos grampositivos, la opsonización mediada por complemento es más importante para su eliminación.
Citocinas
Las citocinas son proteínas solubles que actúan como señales de comunicación entre las células. En una respuesta inmune innata inespecífica, se pueden liberar diversas citocinas para estimular la producción de mediadores químicos u otras funciones celulares, como la proliferación celular, la diferenciación celular, la inhibición de la división celular, la apoptosis y la quimiotaxis.
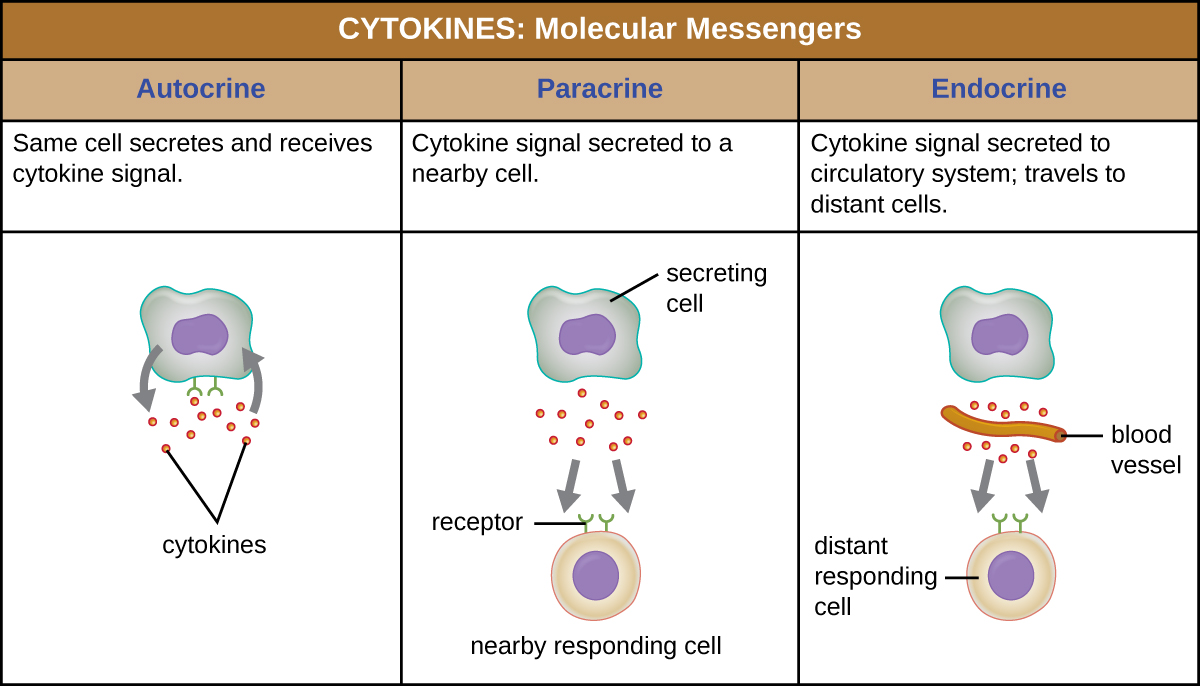
Cuando una citocina se une a su receptor diana, el efecto puede variar ampliamente dependiendo del tipo de citocina y del tipo de célula o receptor al que se haya unido. La función de una citocina particular puede describirse como autocrina, paracrina o endocrina (Figura\(\PageIndex{3}\)). En función autocrina, la misma célula que libera la citocina es la receptora de la señal; en otras palabras, la función autocrina es una forma de autoestimulación por parte de una célula. En contraste, la función paracrina implica la liberación de citocinas de una célula a otras células cercanas, estimulando alguna respuesta de las células receptoras. Por último, la función endocrina ocurre cuando las células liberan citocinas en el torrente sanguíneo para ser transportadas a las células diana mucho más alejadas.
Tres clases importantes de citoquinas son las interleucinas, quimiocinas e interferones. Originalmente se pensaba que las interleucinas eran producidas únicamente por leucocitos (glóbulos blancos) y que solo estimulaban los leucocitos, de ahí las razones de su nombre. Aunque las interleucinas están involucradas en la modulación de casi todas las funciones del sistema inmune, su papel en el cuerpo no se restringe a la inmunidad. Las interleucinas también son producidas por y estimulan una variedad de células no relacionadas con las defensas inmunitarias.
Las quimiocinas son factores quimiotácticos que reclutan leucocitos a sitios de infección, daño tisular e inflamación. A diferencia de factores quimiotácticos más generales, como el factor del complemento C5a, las quimiocinas son muy específicas en los subconjuntos de leucocitos que reclutan.
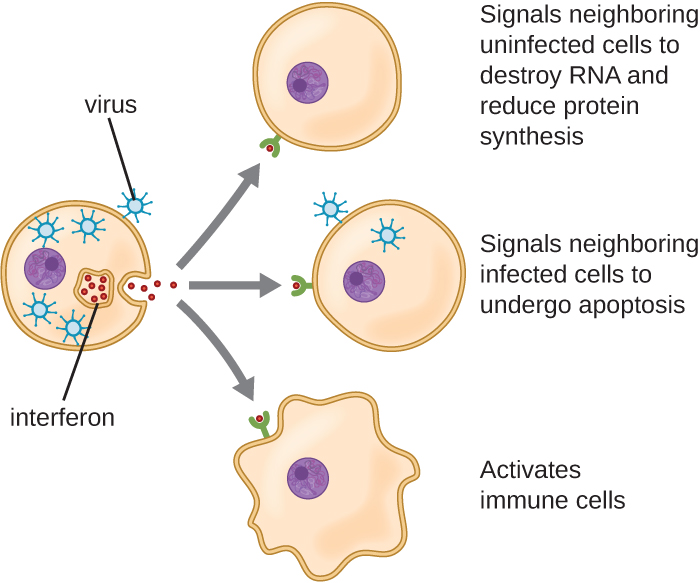
Los interferones son un grupo diverso de moléculas de señalización inmune y son especialmente importantes en nuestra defensa contra los virus. Los interferones tipo I (interferón-α e interferón-β) son producidos y liberados por las células infectadas con virus. Estos interferones estimulan a las células cercanas para detener la producción de ARNm, destruir el ARN ya producido y reducir la síntesis de proteínas. Estos cambios celulares inhiben la replicación viral y la producción de virus maduros, ralentizando la propagación del virus. Los interferones tipo I también estimulan diversas células inmunitarias involucradas en el aclaramiento viral para atacar de manera más agresiva a las células infectadas por virus. El interferón tipo II (interferón-γ) es un activador importante de las células inmunes (Figura\(\PageIndex{4}\)).
Mediadores que provocan inflamación
Muchos de los mediadores químicos discutidos en esta sección contribuyen de alguna manera a la inflamación y la fiebre, que son respuestas inmunes inespecíficas discutidas con más detalle en Inflamación y Fiebre. Las citocinas estimulan la producción de proteínas de fase aguda como la proteína C reactiva y la lectina de unión a manosa en el hígado. Estas proteínas de fase aguda actúan como opsoninas, activando cascadas de complemento a través de la vía lectina.
Algunas citocinas también se unen a mastocitos y basófilos, induciéndolos a liberar histamina, un compuesto proinflamatorio. Los receptores de histamina se encuentran en una variedad de células y median eventos proinflamatorios, como bronconstricción (endurecimiento de las vías respiratorias) y contracción del músculo liso.
Además de la histamina, los mastocitos pueden liberar otros mediadores químicos, como los leucotrienos. Los leucotrienos son mediadores proinflamatorios basados en lípidos que se producen a partir del metabolismo del ácido araquidónico en la membrana celular de leucocitos y células tisulares. En comparación con los efectos proinflamatorios de la histamina, los de los leucotrienos son más potentes y de mayor duración. Juntos, estos mediadores químicos pueden inducir tos, vómitos y diarrea, que sirven para expulsar patógenos del cuerpo.
Ciertas citoquinas también estimulan la producción de prostaglandinas, mediadores químicos que promueven los efectos inflamatorios de quininas e histaminas. Las prostaglandinas también pueden ayudar a elevar la temperatura corporal, conduciendo a fiebre, lo que promueve las actividades de los glóbulos blancos e inhibe ligeramente el crecimiento de microbios patógenos (ver Inflamación y Fiebre).
Otro mediador inflamatorio, la bradiquinina, contribuye al edema, que ocurre cuando los líquidos y leucocitos se filtran fuera del torrente sanguíneo y hacia los tejidos. Se une a los receptores en las células de las paredes capilares, haciendo que los capilares se dilaten y se vuelvan más permeables a los fluidos.
Ejercicio\(\PageIndex{3}\)
- ¿Qué tienen en común las tres vías de activación del complemento?
- Explicar las señales autocrinas, paracrinas y endocrinas.
- Nombrar dos importantes mediadores inductores de inflamación.
Enfoque Clínico: Parte 2
Para aliviar la constricción de sus vías respiratorias, Angela es tratada inmediatamente con antihistamínicos y se le administran corticosteroides a través de un inhalador, para luego monitorearse por un periodo de tiempo. Aunque su condición no empeora, las drogas no parecen estar aliviando su condición. Ella es ingresada en el hospital para mayor observación, pruebas y tratamiento.
Después del ingreso, un médico realiza pruebas de alergia para tratar de determinar si algo en su entorno podría estar desencadenando una respuesta inflamatoria alérgica. Un médico ordena análisis de sangre para verificar los niveles de citocinas particulares. También se toma una muestra de esputo y se envía al laboratorio para tinción microbiana, cultivo e identificación de patógenos que podrían estar causando una infección.
Ejercicio\(\PageIndex{4}\)
- ¿Qué aspectos del sistema inmunitario innato podrían estar contribuyendo a la constricción de las vías respiratorias de Angela?
- ¿Por qué Angela fue tratada con antihistamínicos?
- ¿Por qué el médico estaría interesado en los niveles de citocinas en la sangre de Ángela?
\(\PageIndex{3}\)La tabla proporciona un resumen de las defensas químicas discutidas en esta sección.
| Defensa | Ejemplos | Función |
|---|---|---|
| Químicos y enzimas en los fluidos corporales | Sebo de las glándulas sebáceas | Proporciona barrera de aceite que protege los poros del folículo piloso de patógenos |
| Ácido oleico de sebo y microbiota cutánea | Disminuye el pH para inhibir patógenos | |
| Lisozima en secreciones | Mata bacterias atacando la pared celular | |
| Ácido en el estómago, la orina y la vagina | Inhibe o mata bacterias | |
| Enzimas digestivas y bilis | Matar bacterias | |
| Lactoferrina y transferrina | Enlazar y secuestrar hierro, inhibiendo el crecimiento bacteriano | |
| Surfactante en pulmones | Mata bacterias | |
| Péptidos antimicrobianos | Defensinas, bacteriocinas, dermicidina, catelicidina, histatinas, | Mata bacterias atacando membranas o interfiriendo con las funciones celulares |
| Mediadores de proteínas plasmáticas | Proteínas de fase aguda (proteína C reactiva, amiloide A sérico, ferritina, fibrinógeno, transferrina y lectina de unión a manosa) | Inhibir el crecimiento de bacterias y ayudar a atrapar y matar bacterias |
| Complementos C3b y C4b | Opsonización de patógenos para ayudar a la fagocitosis | |
| Complemento C5a | Quimioatrayente para fagocitos | |
| Complementos C3a y C5a | Anafilatoxinas proinflamatorias | |
| Citocinas | Interleucinas | Estimular y modular la mayoría de las funciones del sistema inmunológico |
| Quimiocinas | Reclutar glóbulos blancos al área infectada | |
| Interferones | Alertar a las células de infección viral, inducir apoptosis de células infectadas por virus, inducir defensas antivirales en células infectadas y no infectadas cercanas, estimular las células inmunitarias para atacar a las células infectadas por virus | |
| Mediadores que provocan inflamación | Histamina | Promueve la vasodilatación, broncoconstricción, contracción del músculo liso, aumento de la secreción y producción de moco |
| Leucotrienos | Promueve la inflamación; más fuerte y duradero que la histamina | |
| Prostaglandinas | Promueve la inflamación y la fiebre | |
| Bradiquinina | Aumenta la vasodilatación y la permeabilidad vascular, lo que lleva a edema |
Conceptos clave y resumen
- Numerosos mediadores químicos producidos endógenamente y exógenamente exhiben funciones antimicrobianas inespecíficas.
- Muchos mediadores químicos se encuentran en fluidos corporales como sebo, saliva, moco, fluidos gástricos e intestinales, orina, lágrimas, cerumen y secreciones vaginales.
- Los péptidos antimicrobianos (AMP) que se encuentran en la piel y en otras áreas del cuerpo se producen en gran medida en respuesta a la presencia de patógenos. Estos incluyen dermcidina, catelicidina, defensinas, histatinas y bacteriocinas.
- El plasma contiene varias proteínas que sirven como mediadores químicos, incluyendo proteínas de fase aguda, proteínas del complemento y citoquinas.
- El sistema del complemento involucra numerosas proteínas precursoras que circulan en plasma. Estas proteínas se activan en una secuencia en cascada en presencia de microbios, dando como resultado la opsonización de patógenos, quimioatracción de leucocitos, inducción de inflamación y citólisis a través de la formación de un complejo de ataque a la membrana (MAC).
- Las citocinas son proteínas que facilitan diversas respuestas inespecíficas por parte de las células inmunitarias innatas, incluyendo la producción de otros mediadores químicos, la proliferación celular, la muerte celular y la diferenciación.
- Las citocinas juegan un papel clave en la respuesta inflamatoria, desencadenando la producción de mediadores inductores de inflamación como proteínas de fase aguda, histamina, leucotrienos, prostaglandinas y bradiquinina.