
1.4: Vías Metabólicas Básicas
- Page ID
- 77447

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Resumen del Metabolismo Básico
- La glucólisis (citosol) y el ciclo de TCA (mitocondrias) convierten la glucosa en moléculas de alta energía: ________ y ___________ y __________.
Esto es solo el comienzo de la producción de energía. NADH y FADH 2 se pueden convertir a más ATP. La fosforilación oxidativa es una vía metabólica que transfiere energía del NADH a la síntesis de ATP en las mitocondrias.
- La oxidación del NADH ocurre en muchos pasos. ¿Por qué las células no hacen esta reacción directamente? (Pista: ¡Esta es una reacción de hidruro!)
Ubicaciones celulares
Los electrones almacenados en forma de las coenzimas reducidas, NADH o FADH 2, se pasan a través de una cadena de proteínas y coenzimas para reducir O 2 —el aceptor de electrones terminal— en H 2 O.
- El NADH se forma en qué punto del metabolismo: ___________.
- El ciclo de TCA ocurre en __________.
- Esta transferencia electrónica de fosforilación oxidativa ocurre en ________________.
Producción de ATP
La energía liberada por los electrones que fluyen a través de esta cadena de transporte de electrones se utiliza para transportar protones para generar un gradiente de pH a través de la membrana.
- La fosforilación de ADP para formar ATP es [endotérmica o exotérmica].
- Los protones fluyen de nuevo a través de la membrana para restaurar el equilibrio. Este proceso es [difusión o transporte activo] y puede impulsar una reacción.
Metabolismo Básico: Glicolisis
La glucosa se metaboliza para producir energía (ATP) para la célula con la liberación de CO 2 y H 2 O como subproductos. La glucólisis es una serie de reacciones catalizadas por enzimas que rompen la glucosa en 2 equivalentes de piruvato. Este proceso (resumido a continuación) también se llama la vía Embden-Meyerhoff.
- ¿Cuántos ATP se producen en este proceso? Ten en cuenta que todo se duplica después de que la glucosa a 6 C se escinde en 2 unidades 3C.
- ¿Cuántos ATP se consumen?
- La glucólisis da como resultado la formación neta de:
- ______ NADH
- ______ ATP
- ______ H 2 O
- ¿La glucólisis es un proceso cuesta arriba o cuesta abajo? (es decir, ¿exotérmica o endotérmica?)
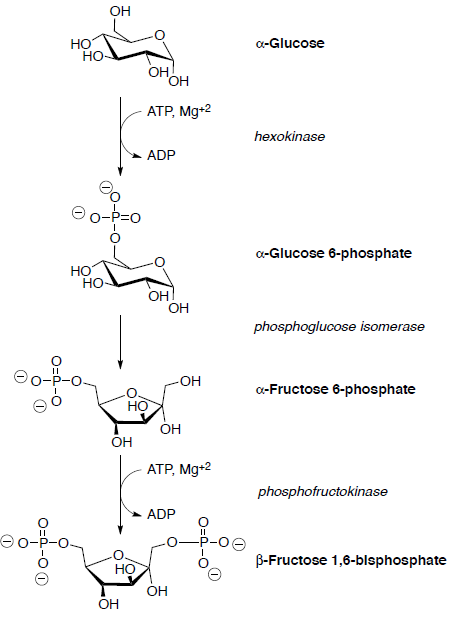
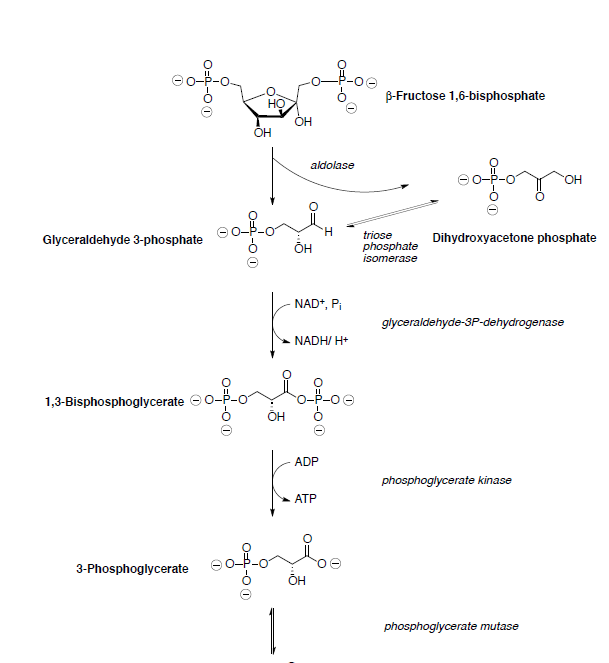
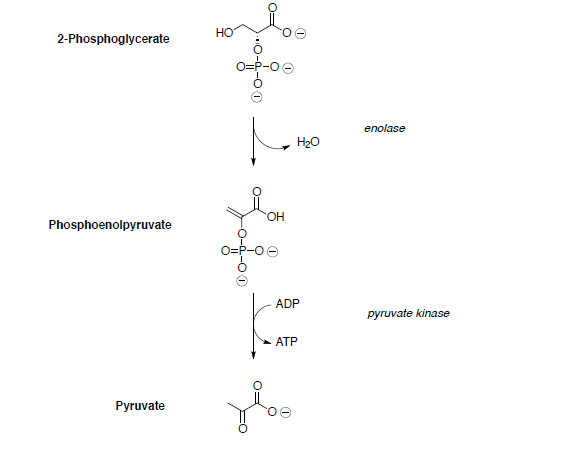
Supongamos que todas las reacciones tienen lugar dentro de una enzima.
La glucosa se fosforila primero en el grupo hidroxilo en C6 por reacción con ATP.
- Proponer un mecanismo para esta reacción.
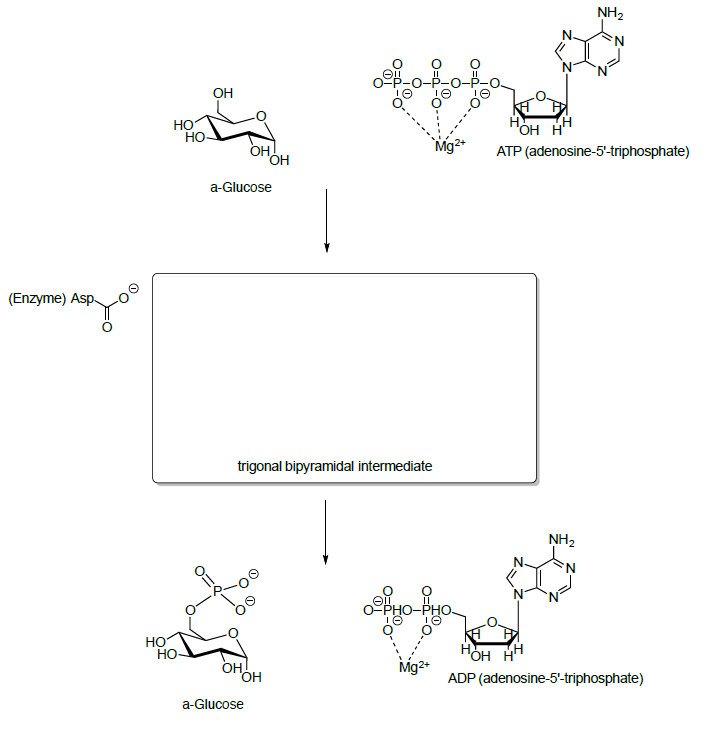
- El ATP no es tan reactivo por sí solo. ¿Por qué?
- Explique por qué un éster de fosfato es un buen electrófilo cuando el Mg +2 está alrededor.
La glucosa-6-fosfato se isomeriza a fructosa-6-fosfato en la siguiente etapa. La interconversión glucosa-fructosa es un proceso multietapa cuyos detalles aún no se entienden completamente.
Comienza con la apertura del hemiacetal a un aldehído de cadena abierta.
- Proponer un mecanismo para esta reacción.
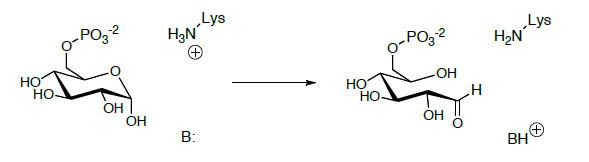
El aldehído de cadena abierta se somete a tautomerización de ceto-enol al enediol que se tautomeriza adicionalmente a una forma ceto-enol diferente.
- Mirando las estructuras de los azúcares, proponer un mecanismo para esta reacción.
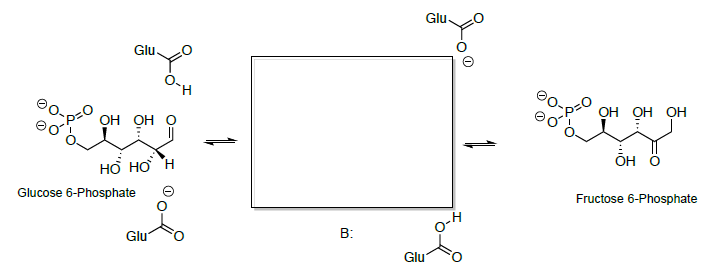
La ciclación de la hidroxicetona de cadena abierta da fructosa (hemiacetal).
- Show a mechanism.
- Predict the product.

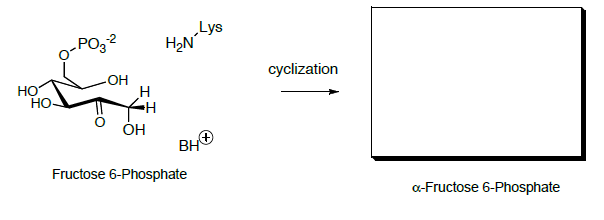
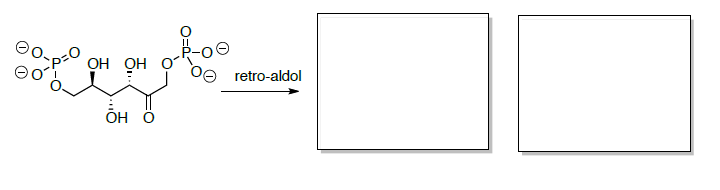
La fructosa-6-fosfato se convierte luego en fructosa 1,6-bisfosfato que posteriormente se escinde en dos compuestos de tres carbonos a través de un retro-aldol.
Crítica: reacción aldol
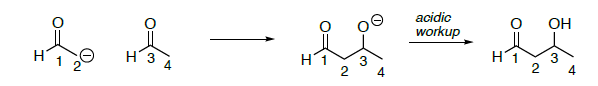
- Sobre la reacción aldol anterior,
- Poner un círculo alrededor del nucleófilo
- Pon una caja alrededor del electrófilo en tus materiales de partida
- Resaltar el enlace que se forma (roto en la reacción retro)
Retro-Aldol
Si la reacción se conduce a materiales de partida (retro-aldol), entonces la reacción favorecerá los materiales de partida.
- Dibujar el mecanismo para el retro-aldol al comenzar con fructosa 1,6-bisfosfato.
- Predecir los dos productos de esta reacción retro aldólica.
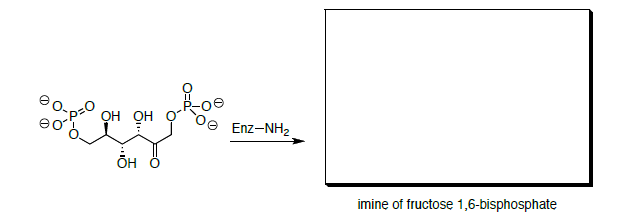
Este mecanismo en realidad se completa con una imine. La fructosa 1,6-bisfosfato reacciona primero con el grupo amino de un residuo de lisina de una enzima.
- Dibujar un mecanismo para la formación de la imina.
La imina puede entonces hacer una reacción de 'retro-cigüeña enamina' (similar al retro-aldol).
Revisión: Stork Enamine (un adol con la enamina reemplazando al anión enolato como nucleófilo).
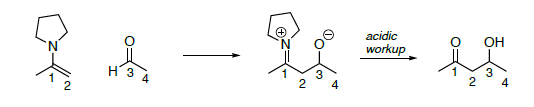
- Sobre la reacción de enamina anterior,
- Poner un círculo alrededor del nucleófilo
- Pon una caja alrededor del electrófilo en tus materiales de partida
- Resaltar el enlace que se forma (roto en la reacción retro)
Enamina Retro-Stork
Si la reacción es conducida a los materiales de partida (retro-cigüeña enamina), entonces la reacción favorecerá los materiales de partida enamina y aldol.
- Predecir los dos productos formados.
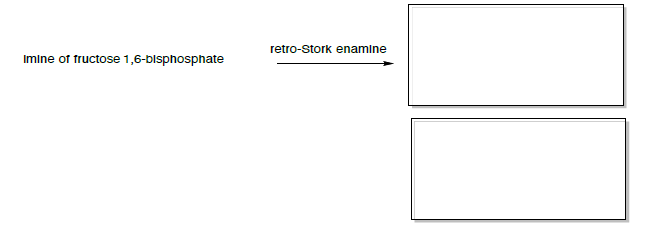
Los productos de la enamina Retro-cigüeña son la enamina de fosfato de dihidroxiacetona y 3-fosfato de gliceraldehído (se muestra a continuación).
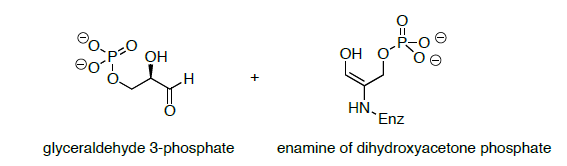
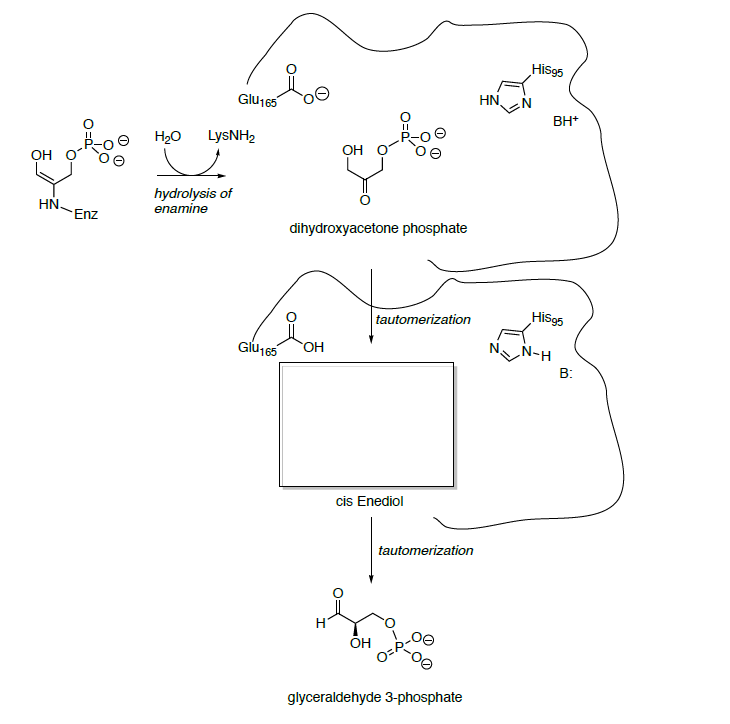
- Propose a mechanism for the conversion of the enamine of dihydroxyacetone phosphate is converted to a second molecule of glyceraldehyde 3-phosphate.
Glyceraldehyde 3-phosphate is oxidized and phosphorylated to 1,3-bisphosphoglycerate.
- Show the mechanisms for this transformation.
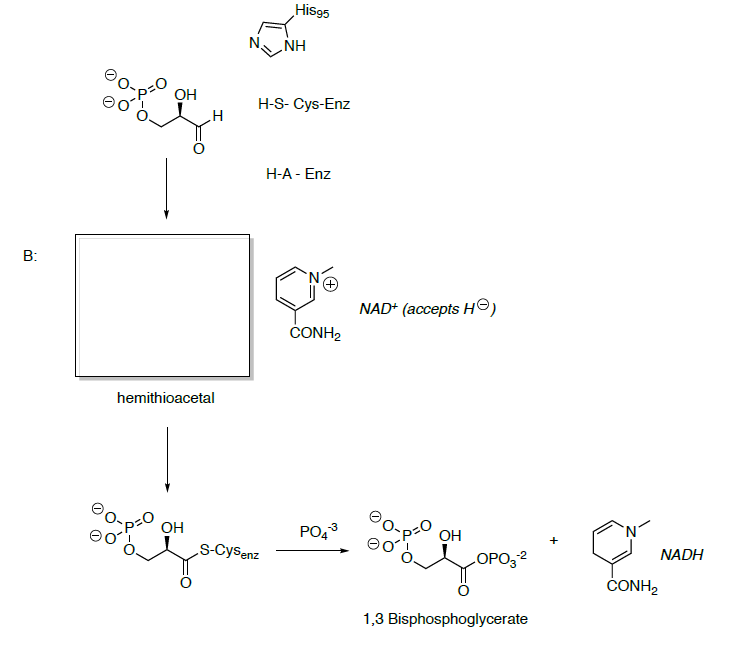
- What is the functional group formed in 1,3-bisphosphoglycerate?
- Predict the reactivity of this carbonyl.
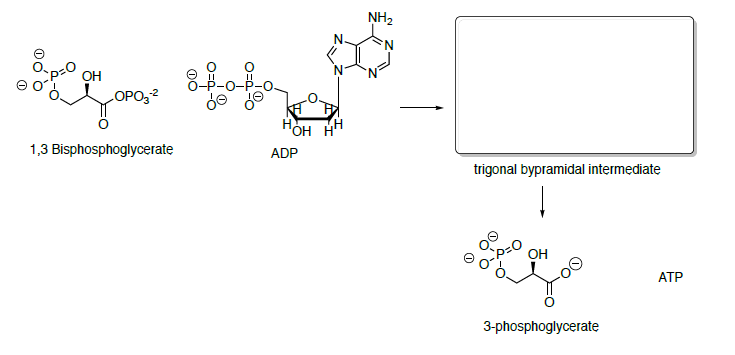
Phosphoglycerate kinase catalyzes the transfer of a phosphoryl group from 1,3-bisphosphoglycerate to ADP forming ATP and 3-phosphoglycerate.
- Propose a mechanism for this transformation.
3-phosphoglycerate is converted to phosphoenol pyruvate (PEP) through dehydration and dephosphorylation.
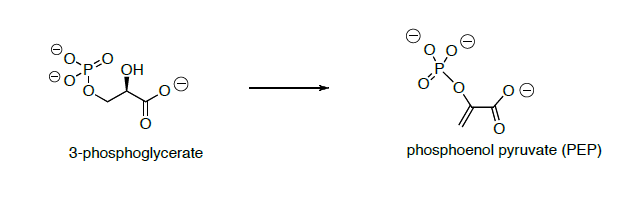
In the last step of the metabolic breakdown of sugars (glycolysis), an enol phosphate is converted to pyruvic acid (shown below). The pyruvic acid is then converted to Acetyl Co A, which is the beginning of the TCA cycle.
- Draw a mechanism for the conversion of the enol phosphate to pyruvic acid.
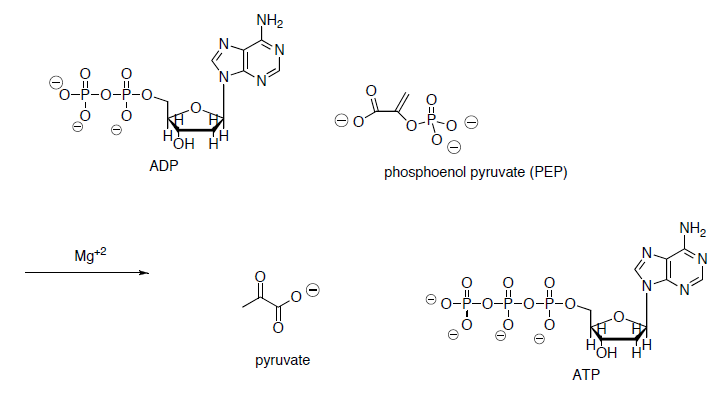
- What drives this reaction? (ie what factors make this reaction energetically favorable?)
Basic Metabolism: TCA Cycle
Hans Krebs and Fritz Lipmann shared the Nobel Prize for Physiology and Medicine in 1953 for their work on elucidating the Krebs cycle and coenzyme A. The Krebs Cycle [or tricarboxylic acid (TCA) or citric acid cycle] plays a central role in the metabolism of glucose to produce energy (ATP). The TCA cycle results ultimately in the oxidation of acetic acid to two molecules of carbon dioxide.
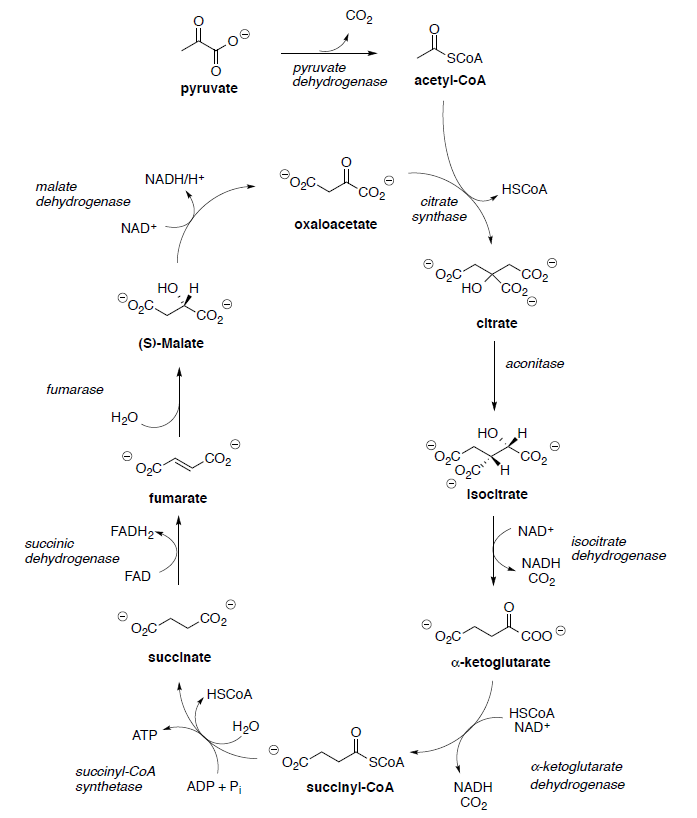
Pyruvate (end product of glycolysis) must be converted to acetyl CoA to enter the TCA cycle.
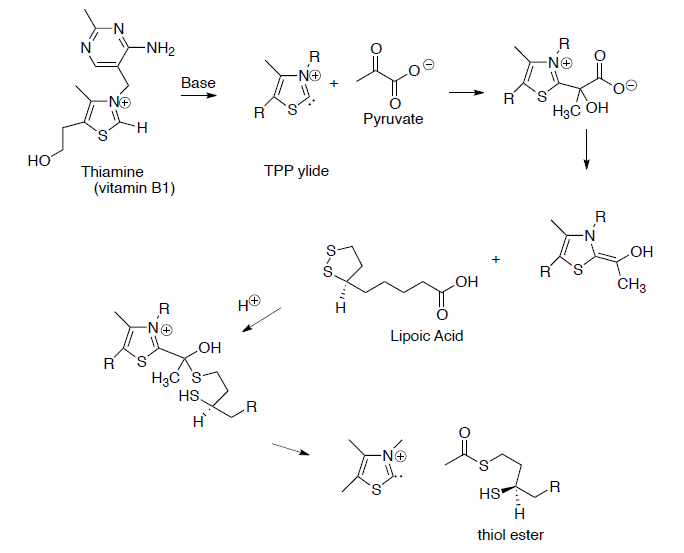
This process begins with the formation of a thiol ester from pyruvate.
- Draw reaction mechanisms for the steps shown below.
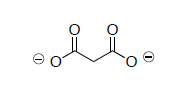
- In his experiments that led to the elucidation of the TCA cycle, Hans Krebs added malonate (shown below) to extracts of pigeon flight muscle. The malonate could not be used as a substrate to replace pyruvate in the pathway above. Why can’t malonate be used? (Think of the carbonyl hill).
At this point, co-enzyme A reacts with the thiol ester (formed in question on previous page) to form acetyl CoA (shown below). To help keep track of the sulfurs, one is in a box and one is in a circle.
- Draw the mechanism for this reaction.
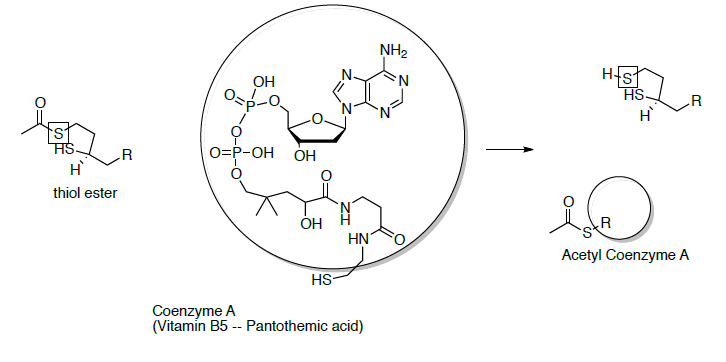
- The thiol ester formed in the last step of the reaction above is an ‘activated carbonyl’ (i.e. a better electrophile). Explain why the thiol ester is a better electrophile than the carboxylate anion.
- In an equivalent organic chemistry reaction, what would you use as an ‘activated carbonyl’?
In the next step Acetyl CoA reacts with oxaloacetate to form citryl CoA.
- Propose a mechanism for this reaction.
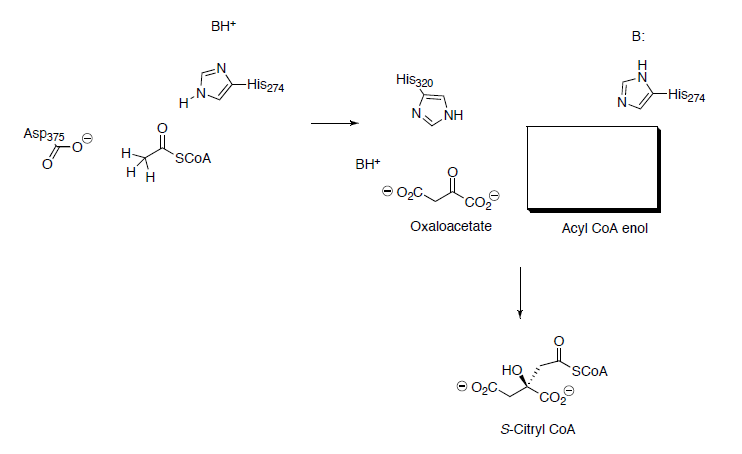
- In a similar reaction in organic chemistry, what would be the product for the reaction below? What type of reaction is this?
Citryl CoA is then hydrolyzed to citrate.
- Propose a reaction mechanism for this reaction.
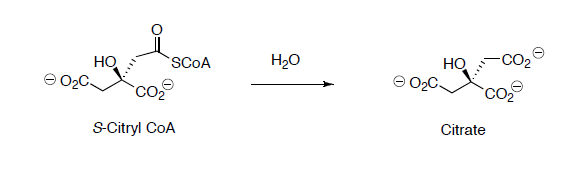
Citrate is converted to isocitrate through two steps.
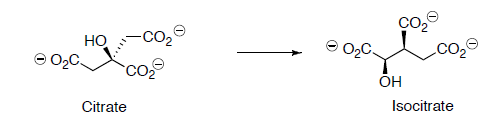
- Label all chiral centers with R or S.
- What changed in the conversion of citrate to isocitrate?
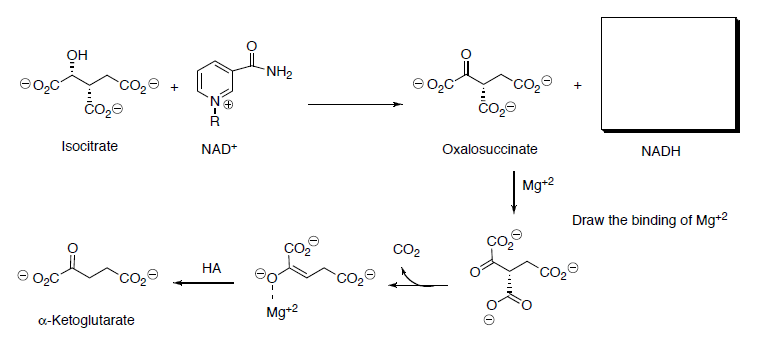
Isocitrate is oxidized to oxalosuccinate with NAD+.
- Draw the mechanism (and the other product) for this reaction.
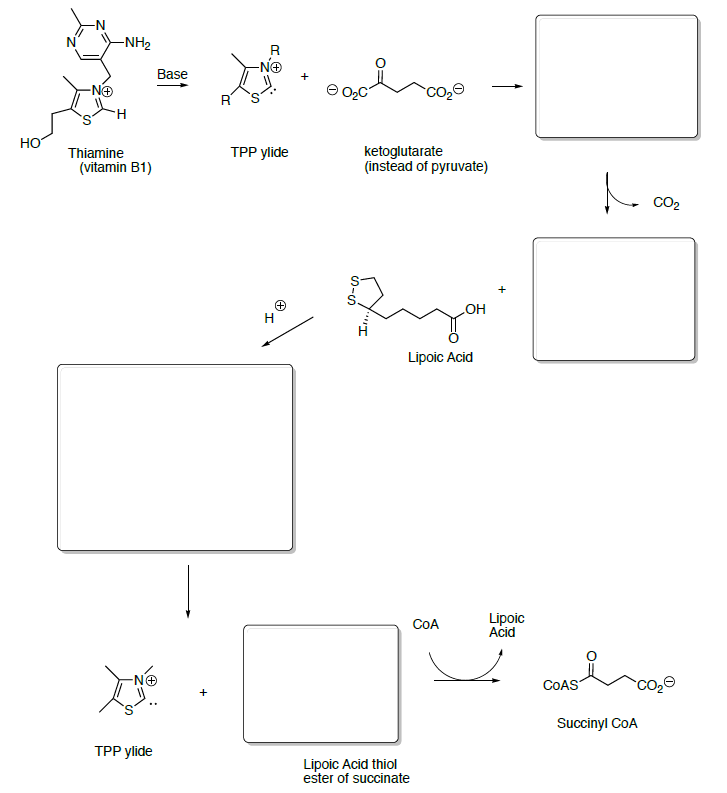
Ketoglutarate is transformed to succinyl CoA in a multistep process analogous to the transformation of pyruvate to acetyl CoA that we saw in the first step.
- Draw the transformation starting with the reaction with TPP ylide.
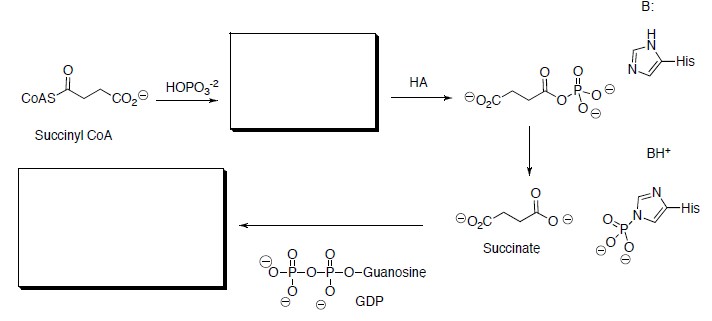
Succinyl CoA is hydrolyzed to succinate and is coupled with the phosphorylation of guanosine diphosphate (GDP) to give guanosine triphosphate (GTP).
- Draw the mechanism for this reaction.
Basic Metabolism: Oxidative Phosphorylation
Electron Transfer in Complex I
Complex I is located in the inner mitochondrial membrane in eukaryotes. The electrons from NADH (produced in the TCA cycle) begin to be shuttled through small steps to capture the energy.
This section will examine the mechanisms of electron transfer by the peripheral domain, proton transfer by the membrane domain and how their coupling can drive proton transport.
The net reaction of Complex I is the oxidation of NADH and the reduction of ubiquinone.
Net reaction:
\[\ce{NADH + H^+ + UQ \rightarrow NAD^+ + UQH2}\]
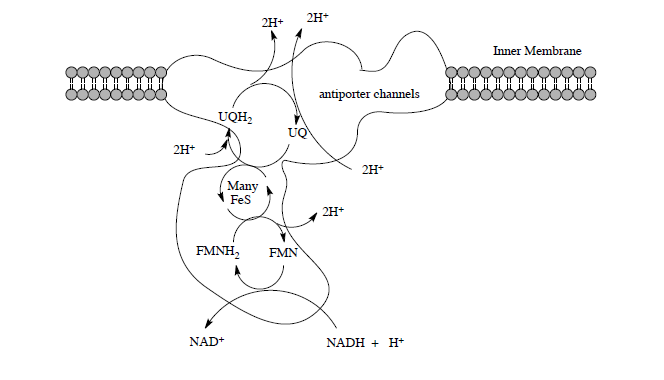
- How many protons are moved across the membrane for each cycle of Complex I?
- Is this active transport or passive diffusion?
- If this is active, what is fueling this transport?
- Is this with or against the concentration gradient? (i.e. antiporter or synporter?)
Complex II: Overview
Complex II (aka succinate dehydrogenase from the TCA cycle) oxidizes succinate (–O2CCH2CH2CO2–) to fumarate (trans-–O2CCH=CHCO2–).
Complex II also has a cascade of electron transfers. When succinate is converted to fumarate, the electrons are passed through a new cascade to eventually reduce UQ (just like Complex I!)
\[\ce{succinate \rightarrow fumarate + 2H+ + 2e-}\]
\[\ce{UQ + 2H+ + 2e- \rightarrow UQH2}\]
- Write the net reaction for the work of Complex II.
- The reaction catalyzed in Complex II has a very small ΔG°. Is it sufficient to power an antiporter channel?
Complex III: Overview
Complex III (sometimes called cytochrome bc1 complex) has two main substrates: cytochrome c and UQH2. The structure of this complex was determined by Johann Deisenhofer (Nobel Prize for a photosynthetic reaction center – we will see this soon).
This role of complex III is to transfer the electrons from UQH2 to cytochrome c.
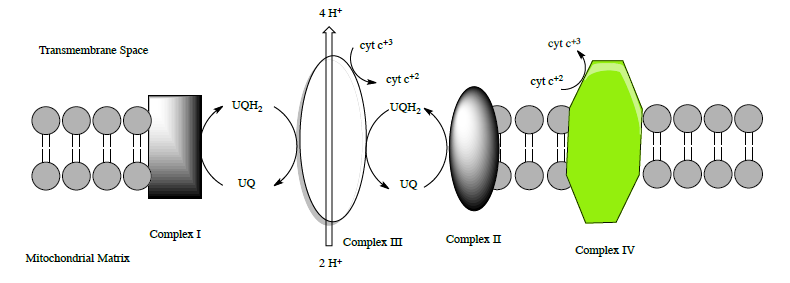
- Complete the equation for the redox reactions of complex III.
___ UQH2 + 1 UQ + 2 H+ + ___ cyt c+3 \(\ce{\rightarrow}\) ___ UQH2 + ___ UQ + 4 H+ + ___ cyt c+2
• There are two H+ coming from the mitochondrial matrix but _____ H+ are transported into the
inter-membrane space
Complex III to Complex IV: Cytochrome C as a mobile carrier
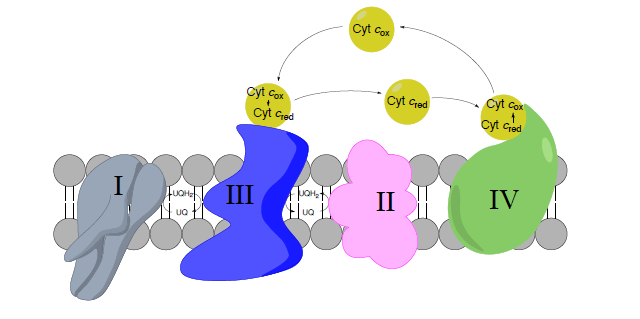
- Encierra en círculo los portadores de electrones móviles en la imagen de arriba.
Descripción general del complejo IV
¡Otro complejo cuyo objetivo es mover electrones y protones! Este es el gran paso ya que es el sitio principal para la
utilización de dioxígeno en todos los organismos anaerobios. La estructura del complejo IV se muestra en la figura izquierda y a la derecha en un diagrama tomado de las vías de Kegg (con permiso).
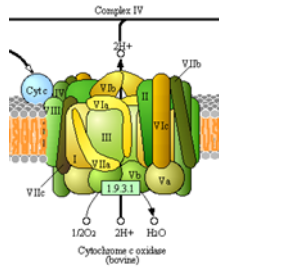
- Completar la ecuación neta para las reacciones redox del complejo IV.
___ cyt c +2 + 1 O 2 + 8 H +\(\ce{\rightarrow}\) ___ H 2 O + 4 H + + ___ cyt c +3
- ¿Cuántos protones están siendo “bombeados” en el espacio intermembrana? _________
- ¿Cuántos electrones se necesitan para equilibrar esta ecuación? _________
- ¿Cuáles son los portadores “móviles” iniciales y finales de electrones?
Complejo V: ATP Synthase
Despreciando el Complejo II, la reacción general de la cadena mitocondrial, por 2e — transferida, puede escribirse como:
\[\ce{NADH + H+ + ½ O2 + 10 H+("in") \rightarrow NAD+ + H2O + 10 H+("out")} E° = +1.135V\]
Cada dos e — (de 1 molécula de NADH) a través de la cadena de transporte de electrones da como resultado la transferencia neta de 10 protones a través de la membrana:
- Complejo I: ________ H +
- Complejo III: ________ H +
- Complejo IV: ________ H +
Los protones se difundirán de un área de alta concentración de protones a un área de menor concentración de protones. Peter Mitchell recibió el Premio Nobel en 1978 por su propuesta de que un gradiente de concentración
electroquímica de protones a través de una membrana podría aprovecharse para hacer ATP. El gradiente de protones creado por la cadena de transporte de electrones proporciona suficiente energía para sintetizar aproximadamente 2.5 moléculas de ATP a través de un proceso llamado quimiósmosis.
- Este flujo de protones es impulsado por dos fuerzas (rellene los espacios en blanco):
- Fuerza de difusión causada por un gradiente de concentración. Todas las partículas tienden a pasar de __________ concentración a __________ concentración.
- Fuerza electrostática causada por un gradiente de potencial eléctrico. Un gradiente eléctrico es consecuencia de la separación de carga. Los protones se sentirán atraídos por ___________.
La ATP sintasa es una enzima importante que utiliza el gradiente de protones para impulsar la síntesis de (ATP).
Motor de impulsiones de potencial eléctrico
El rotor no está bloqueado en una posición fija en el centro de la bicapa y los sitios del rotor cambian entre los estados vacío y unido a iones. Al impulsar la síntesis de ATP, un ion llega del periplasma y se une a un sitio de rotor vacío.
La carga positiva del estator (Arg 227) juega un papel fundamental en la función del motor F 0.
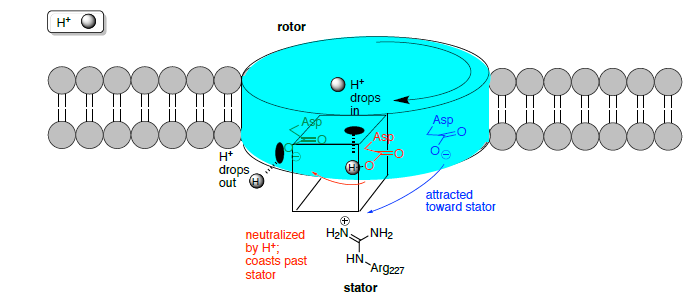
- Cuál es el cargo de los sitios de encuadernación vacíos:
- cuando no hay iones ligados?
- cuando un ion Na + /H + se une?
- Cuando un ion ingresa al sitio del rotor desde el canal del estator, la carga neta se reduce, por lo tanto [aumenta/disminuye] la atracción hacia el estator. Ahora el rotor es capaz de moverse a través de la parte hidrofóbica del estator, mientras que la arginina atrae al siguiente sitio vacío del rotor.
- El sitio vacío (carga = ________) es atraído electrostáticamente por el estator (carga = ________) y guiado a la siguiente ranura.
- El sitio del rotor está ocupado hasta que alcanza el estator desde el lado opuesto, donde encuentra la carga positiva del estator, provocando la disociación del ion. ¿Por qué? Considera gradientes de difusión y cargas.
Energía Eléctrica Combustibles Rotativos ATP Sintasa
- Rellene los espacios en blanco del siguiente resumen de ATP Sintasa:
Durante la síntesis de ATP, el gradiente ____________ alimenta el motor F 0 incrustado en la membrana para rotar el tallo central. Esta rotación provoca cambios de unión secuenciales en el dominio F 1 periférico de manera que un sitio catalítico se une a ________ y fosfato, el segundo hace ATP fuertemente unido, y el tercer paso ____________.
En bacterias de crecimiento anaeróbico, cuando las enzimas respiratorias no están activas, el motor F 1 puede hidrolizar ATP.
- ¿En qué dirección girará la bomba en estas condiciones?
- ¿Qué pasará con el motor F 0? ¿Y el gradiente H +?
Fuentes
Dimroth, Operación del motor F0 de la ATP sintasa, Biochimica et Biophysica Acta (BBA) -Bioenergetics, 2000, 1458, 374-386.