
21.2: Defensas de barrera y respuesta inmune innata
- Page ID
- 123205

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Describir las defensas de barrera del cuerpo
- Mostrar cómo es importante la respuesta inmune innata y cómo ayuda a guiar y preparar al cuerpo para las respuestas inmunitarias adaptativas
- Describir diversos factores solubles que forman parte de la respuesta inmune innata
- Explicar los pasos de la inflamación y cómo conducen a la destrucción de un patógeno
- Discutir las respuestas inmunes inducidas tempranas y su nivel de efectividad
El sistema inmune se puede dividir en dos mecanismos superpuestos para destruir patógenos: la respuesta inmune innata, que es relativamente rápida pero inespecífica y por lo tanto no siempre efectiva, y la respuesta inmune adaptativa, que es más lenta en su desarrollo durante una infección inicial con un patógeno, pero es altamente específicos y efectivos para atacar una amplia variedad de patógenos (Figura\(\PageIndex{1}\)).
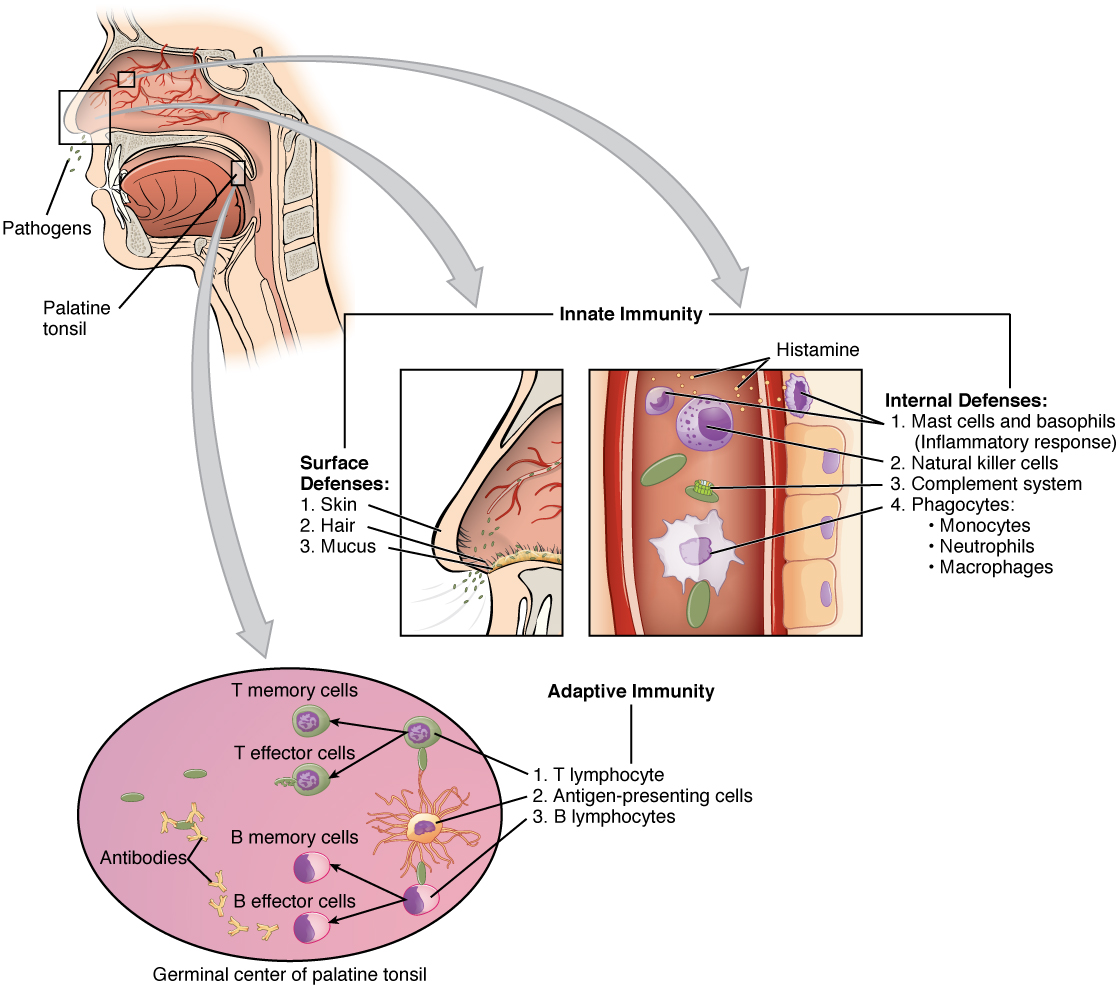
Cualquier discusión sobre la respuesta inmune innata suele comenzar con las barreras físicas que impiden que los patógenos ingresen al cuerpo, los destruyen después de que ingresan, o los eliminan antes de que puedan establecerse en el ambiente hospitalario de los tejidos blandos del cuerpo. Las defensas de barrera forman parte de los mecanismos de defensa más básicos del organismo. Las defensas de barrera no son una respuesta a las infecciones, pero están trabajando continuamente para proteger contra una amplia gama de patógenos.
Los diferentes modos de defensas de barrera están asociados con las superficies externas del organismo, donde los patógenos pueden intentar ingresar (Tabla). La principal barrera para la entrada de microorganismos en el cuerpo es la piel. No sólo la piel está cubierta con una capa de epitelio muerto y queratinizado que es demasiado seco para que las bacterias crezcan, sino que como estas células se desprenden continuamente de la piel, transportan bacterias y otros patógenos con ellas. Además, el sudor y otras secreciones de la piel pueden disminuir el pH, contener lípidos tóxicos y lavar físicamente los microbios.
| Defensas de Barrera | ||
|---|---|---|
| Sitio | Defensa específica | Aspecto protector |
| Piel | Superficie epidérmica | Células queratinizadas de superficie, células de Langerhans |
| Piel (sudor/secreciones) | Glándulas sudoríparas, glándulas sebáceas | pH bajo, acción de lavado |
| Cavidad oral | Glándulas salivales | Lisozima |
| Estómago | Tracto gastrointestinal | pH bajo |
| Superficies mucosas | Epitelio mucoso | Células epiteliales no queratinizadas |
| Flora normal (bacterias no patógenas) | Tejidos mucosos | Evitar que los patógenos crezcan en las superficies mucosas |
Otra barrera es la saliva en la boca, que es rica en lisozima, una enzima que destruye las bacterias al digerir sus paredes celulares. El ambiente ácido del estómago, que es fatal para muchos patógenos, también es una barrera. Adicionalmente, la capa de moco del tracto gastrointestinal, tracto respiratorio, tracto reproductivo, ojos, oídos y nariz atrapa tanto microbios como desechos, y facilita su eliminación. En el caso del tracto respiratorio superior, las células epiteliales ciliadas mueven el moco potencialmente contaminado hacia arriba hasta la boca, donde luego se ingiere en el tracto digestivo, terminando en el duro ambiente ácido del estómago. Considerando la frecuencia con la que respiras en comparación con la frecuencia con la que comes o realizas otras actividades que te exponen a patógenos, no es de extrañar que hayan evolucionado múltiples mecanismos de barrera para trabajar en concierto para proteger esta área vital.
Células de la Respuesta Inmune Innata
Un fagocito es una célula que es capaz de rodear y engullir una partícula o célula, un proceso llamado fagocitosis. Los fagocitos del sistema inmune engullen otras partículas o células, ya sea para limpiar un área de escombros, células viejas, o para matar organismos patógenos como bacterias. Los fagocitos son la acción rápida del organismo, primera línea de defensa inmunológica contra organismos que han violado las defensas de barrera y han ingresado a los tejidos vulnerables del organismo.
Fagocitos: Macrófagos y Neutrófilos
Muchas de las células del sistema inmune tienen una capacidad fagocítica, al menos en algún momento durante sus ciclos de vida. La fagocitosis es un mecanismo importante y eficaz para destruir patógenos durante las respuestas inmunes innatas. El fagocito toma el organismo dentro de sí mismo como un fagosoma, que posteriormente se fusiona con un lisosoma y sus enzimas digestivas, matando efectivamente a muchos patógenos. Por otro lado, algunas bacterias como la Mycobacteria tuberculosis, la causa de la tuberculosis, pueden ser resistentes a estas enzimas y por lo tanto son mucho más difíciles de eliminar del organismo. Los macrófagos, neutrófilos y células dendríticas son los principales fagocitos del sistema inmune.
Un macrófago es un fagocito de forma irregular que es de naturaleza ameboide y es el fagocitos más versátil del cuerpo. Los macrófagos se mueven a través de los tejidos y aprietan a través de las paredes capilares usando pseudopodios No solo participan en respuestas inmunes innatas sino que también han evolucionado para cooperar con los linfocitos como parte de la respuesta inmune adaptativa. Los macrófagos existen en muchos tejidos del cuerpo, ya sea vagando libremente a través de los tejidos conectivos o fijados a fibras reticulares dentro de tejidos específicos como los ganglios linfáticos. Cuando los patógenos violan las defensas de barrera del organismo, los macrófagos son la primera línea de defensa (Tabla). Se les llama diferentes nombres, dependiendo del tejido: células de Kupffer en el hígado, histiocitos en tejido conectivo y macrófagos alveolares en los pulmones.
Un neutrófilo es una célula fagocítica que es atraída a través de la quimiotaxis desde el torrente sanguíneo hacia los tejidos infectados. Estas células esféricas son granulocitos. Un granulocito contiene gránulos citoplásmicos, los cuales a su vez contienen una variedad de mediadores vasoactivos como la histamina. En contraste, los macrófagos son agranulocitos. Un agranulocito tiene pocos o ningún gránulo citoplásmico. Mientras que los macrófagos actúan como centinelas, siempre en guardia contra la infección, los neutrófilos pueden considerarse como refuerzos militares que son llamados a una batalla para apresurar la destrucción del enemigo. Aunque, generalmente se considera la célula principal que mata patógenos del proceso inflamatorio de la respuesta inmune innata, nuevas investigaciones han sugerido que los neutrófilos también juegan un papel en la respuesta inmune adaptativa, al igual que los macrófagos.
Un monocito es una célula precursora circulante que se diferencia en una célula macrófaga o dendrítica, que puede ser atraída rápidamente a áreas de infección por moléculas señal de inflamación.
| Células Fagocíticas del Sistema Inmune Innato | |||
|---|---|---|---|
| Celda | Tipo de celda | Ubicación primaria | Función en la respuesta inmune innata |
| Macrófagos | Agranulocito | Cavidades corporales/órganos | Fagocitosis |
| Neutrófilos | Granulocito | Sangre | Fagocitosis |
| Monocito | Agranulocito | Sangre | Precursor de macrófagos/células dendríticas |
Células asesinas naturales
Las células NK son un tipo de linfocitos que tienen la capacidad de inducir apoptosis, es decir, muerte celular programada, en células infectadas con patógenos intracelulares como bacterias intracelulares obligadas y virus. Las células NK reconocen estas células por mecanismos que aún no se entienden bien, pero que presumiblemente involucran a sus receptores de superficie. Las células NK pueden inducir apoptosis, en la que una cascada de eventos dentro de la célula provoca su propia muerte por cualquiera de dos mecanismos:
1) Las células NK son capaces de responder a señales químicas y expresar el ligando fas. El ligando fas es una molécula superficial que se une a la molécula fas en la superficie de la célula infectada, enviándole señales apoptóticas, matando así la célula y el patógeno dentro de ella; o
2) Los gránulos de las células NK liberan perforinas y granzimas. Una perforina es una proteína que forma poros en las membranas de las células infectadas. Una granzima es una enzima que digiere proteínas que ingresa a la célula a través de los poros de perforina y desencadena la apoptosis intracelularmente.
Ambos mecanismos son especialmente efectivos contra las células infectadas por virus. Si se induce la apoptosis antes de que el virus tenga la capacidad de sintetizar y ensamblar todos sus componentes, no se liberará ningún virus infeccioso de la célula, evitando así una mayor infección.
Reconocimiento de Patógenos
Las células de la respuesta inmune innata, las células fagocíticas y las células NK citotóxicas reconocen patrones de moléculas específicas de patógenos, como componentes de la pared celular bacteriana o proteínas flagelares bacterianas, utilizando receptores de reconocimiento de patrones. Un receptor de reconocimiento de patrones (PRR) es un receptor unido a membrana que reconoce rasgos característicos de un patógeno y moléculas liberadas por células estresadas o dañadas.
Estos receptores, que se cree que evolucionaron antes de la respuesta inmune adaptativa, están presentes en la superficie celular sean necesarios o no. Su variedad, sin embargo, está limitada por dos factores. En primer lugar, el hecho de que cada tipo de receptor debe ser codificado por un gen específico requiere que la célula asigne la mayor parte o la totalidad de su ADN para hacer receptores capaces de reconocer todos los patógenos. En segundo lugar, la variedad de receptores está limitada por la superficie finita de la membrana celular. Por lo tanto, el sistema inmune innato debe “arreglárselas” utilizando solo un número limitado de receptores que son activos contra una variedad de patógenos tan amplia como sea posible. Esta estrategia está en marcado contraste con el enfoque utilizado por el sistema inmune adaptativo, que utiliza un gran número de receptores diferentes, cada uno altamente específico para un patógeno en particular.
Si las células del sistema inmune innato entran en contacto con una especie de patógeno que reconocen, la célula se unirá al patógeno e iniciará la fagocitosis (o apoptosis celular en el caso de un patógeno intracelular) en un esfuerzo por destruir el microbio infractor. Los receptores varían algo según el tipo de célula, pero suelen incluir receptores para componentes bacterianos y para complemento, que se discuten a continuación.
Mediadores Solubles de la Respuesta Inmune Innata
Las discusiones anteriores han aludido a señales químicas que pueden inducir a las células a cambiar diversas características fisiológicas, como la expresión de un receptor en particular. Estos factores solubles se secretan durante las respuestas innatas o inducidas tempranas, y posteriormente durante las respuestas inmunes adaptativas.
Citocinas y Quimiocinas
Una citocina es una molécula de señalización que permite que las células se comuniquen entre sí a distancias cortas. Las citocinas se secretan al espacio intercelular, y la acción de la citocina induce a la célula receptora a cambiar su fisiología. Una quimiocina es un mediador químico soluble similar a las citocinas excepto que su función es atraer células (quimiotaxis) desde distancias más largas.

Visite este sitio web para conocer la quimiotaxis de fagocitos. La quimiotaxis de fagocitos es el movimiento de los fagocitos según la secreción de mensajeros químicos en forma de interleucinas y otras quimiocinas. ¿Por qué medios un fagocito destruye una bacteria que ha ingerido?
Proteínas tempranas
Las proteínas inducidas tempranas son aquellas que no están constitutivamente presentes en el cuerpo, sino que se elaboran a medida que se necesitan temprano durante la respuesta inmune innata. Los interferones son un ejemplo de proteínas inducidas tempranas. Las células infectadas con virus secretan interferones que viajan a las células adyacentes e inducen a producir proteínas antivirales. Así, a pesar de que se sacrifica la célula inicial, las células circundantes están protegidas. Otras proteínas inducidas tempranas específicas para componentes de la pared celular bacteriana son la proteína de unión a manosa y la proteína C reactiva, elaboradas en el hígado, que se unen específicamente a componentes polisacáridos de la pared celular bacteriana. Los fagocitos como los macrófagos tienen receptores para estas proteínas, y así son capaces de reconocerlas ya que se unen a la bacteria. Esto acerca al fagocito y a la bacteria y mejora la fagocitosis de la bacteria mediante el proceso conocido como opsonización. La opsonización es el etiquetado de un patógeno para la fagocitosis mediante la unión de un anticuerpo o una proteína antimicrobiana.
Sistema Complemento
El sistema del complemento es una serie de proteínas que se encuentran constitutivamente en el plasma sanguíneo. Como tal, estas proteínas no se consideran parte de la respuesta inmune inducida temprana, a pesar de que comparten características con algunas de las proteínas antibacterianas de esta clase. Elaborados en el hígado, tienen una variedad de funciones en la respuesta inmune innata, utilizando lo que se conoce como la “vía alternativa” de activación del complemento. Adicionalmente, el complemento funciona también en la respuesta inmune adaptativa, en lo que se llama la vía clásica. El sistema del complemento consiste en varias proteínas que alteran enzimáticamente y fragmentan posteriormente proteínas en una serie, razón por la cual se denomina cascada. Una vez activada, la serie de reacciones es irreversible, y libera fragmentos que tienen las siguientes acciones:
- Se une a la membrana celular del patógeno que lo activa, marcándolo para fagocitosis (opsonización)
- Difundir lejos del patógeno y actuar como agentes quimiotácticos para atraer las células fagocíticas al sitio de la inflamación
- Formar poros dañinos en la membrana plasmática del patógeno
Figura\(\PageIndex{2}\) shows the classical pathway, which requires antibodies of the adaptive immune response. The alternate pathway does not require an antibody to become activated.

The splitting of the C3 protein is the common step to both pathways. In the alternate pathway, C3 is activated spontaneously and, after reacting with the molecules factor P, factor B, and factor D, splits apart. The larger fragment, C3b, binds to the surface of the pathogen and C3a, the smaller fragment, diffuses outward from the site of activation and attracts phagocytes to the site of infection. Surface-bound C3b then activates the rest of the cascade, with the last five proteins, C5–C9, forming the membrane-attack complex (MAC). The MAC can kill certain pathogens by disrupting their osmotic balance. The MAC is especially effective against a broad range of bacteria. The classical pathway is similar, except the early stages of activation require the presence of antibody bound to antigen, and thus is dependent on the adaptive immune response. The earlier fragments of the cascade also have important functions. Phagocytic cells such as macrophages and neutrophils are attracted to an infection site by chemotactic attraction to smaller complement fragments. Additionally, once they arrive, their receptors for surface-bound C3b opsonize the pathogen for phagocytosis and destruction.
Inflammatory Response
The hallmark of the innate immune response is inflammation. Inflammation is something everyone has experienced. Stub a toe, cut a finger, or do any activity that causes tissue damage and inflammation will result, with its four characteristics: heat, redness, pain, and swelling (“loss of function” is sometimes mentioned as a fifth characteristic). It is important to note that inflammation does not have to be initiated by an infection, but can also be caused by tissue injuries. The release of damaged cellular contents into the site of injury is enough to stimulate the response, even in the absence of breaks in physical barriers that would allow pathogens to enter (by hitting your thumb with a hammer, for example). The inflammatory reaction brings in phagocytic cells to the damaged area to clear cellular debris and to set the stage for wound repair (Figure \(\PageIndex{3}\)).

This reaction also brings in the cells of the innate immune system, allowing them to get rid of the sources of a possible infection. Inflammation is part of a very basic form of immune response. The process not only brings fluid and cells into the site to destroy the pathogen and remove it and debris from the site, but also helps to isolate the site, limiting the spread of the pathogen. Acute inflammation is a short-term inflammatory response to an insult to the body. If the cause of the inflammation is not resolved, however, it can lead to chronic inflammation, which is associated with major tissue destruction and fibrosis. Chronic inflammation is ongoing inflammation. It can be caused by foreign bodies, persistent pathogens, and autoimmune diseases such as rheumatoid arthritis.
There are four important parts to the inflammatory response:
- Tissue Injury. The released contents of injured cells stimulate the release of mast cell granules and their potent inflammatory mediators such as histamine, leukotrienes, and prostaglandins. Histamine increases the diameter of local blood vessels (vasodilation), causing an increase in blood flow. Histamine also increases the permeability of local capillaries, causing plasma to leak out and form interstitial fluid. This causes the swelling associated with inflammation. Additionally, injured cells, phagocytes, and basophils are sources of inflammatory mediators, including prostaglandins and leukotrienes. Leukotrienes attract neutrophils from the blood by chemotaxis and increase vascular permeability. Prostaglandins cause vasodilation by relaxing vascular smooth muscle and are a major cause of the pain associated with inflammation. Nonsteroidal anti-inflammatory drugs such as aspirin and ibuprofen relieve pain by inhibiting prostaglandin production.
- Vasodilation. Many inflammatory mediators such as histamine are vasodilators that increase the diameters of local capillaries. This causes increased blood flow and is responsible for the heat and redness of inflamed tissue. It allows greater access of the blood to the site of inflammation.
- Increased Vascular Permeability. At the same time, inflammatory mediators increase the permeability of the local vasculature, causing leakage of fluid into the interstitial space, resulting in the swelling, or edema, associated with inflammation.
- Recruitment of Phagocytes. Leukotrienes are particularly good at attracting neutrophils from the blood to the site of infection by chemotaxis. Following an early neutrophil infiltrate stimulated by macrophage cytokines, more macrophages are recruited to clean up the debris left over at the site. When local infections are severe, neutrophils are attracted to the sites of infections in large numbers, and as they phagocytose the pathogens and subsequently die, their accumulated cellular remains are visible as pus at the infection site.
Overall, inflammation is valuable for many reasons. Not only are the pathogens killed and debris removed, but the increase in vascular permeability encourages the entry of clotting factors, the first step towards wound repair. Inflammation also facilitates the transport of antigen to lymph nodes by dendritic cells for the development of the adaptive immune response.
Chapter Review
Innate immune responses are critical to the early control of infections. Whereas barrier defenses are the body’s first line of physical defense against pathogens, innate immune responses are the first line of physiological defense. Innate responses occur rapidly, but with less specificity and effectiveness than the adaptive immune response. Innate responses can be caused by a variety of cells, mediators, and antibacterial proteins such as complement. Within the first few days of an infection, another series of antibacterial proteins are induced, each with activities against certain bacteria, including opsonization of certain species. Additionally, interferons are induced that protect cells from viruses in their vicinity. Finally, the innate immune response does not stop when the adaptive immune response is developed. In fact, both can cooperate and one can influence the other in their responses against pathogens.
Interactive Link Questions
Visit this website to learn about phagocyte chemotaxis. Phagocyte chemotaxis is the movement of phagocytes according to the secretion of chemical messengers in the form of interleukins and other chemokines. By what means does a phagocyte destroy a bacterium that it has ingested?
Answer: The bacterium is digested by the phagocyte’s digestive enzymes (contained in its lysosomes).
Review Questions
Q. Which of the following signs is not characteristic of inflammation?
A. redness
B. pain
C. cold
D. swelling
Answer: C
Q. Which of the following is not important in the antiviral innate immune response?
A. interferons
B. natural killer cells
C. complement
D. microphages
Answer: D
Q. Enhanced phagocytosis of a cell by the binding of a specific protein is called ________.
A. endocytosis
B. opsonization
C. anaphylaxis
D. complement activation
Answer: B
Q. Which of the following leads to the redness of inflammation?
A. increased vascular permeability
B. anaphylactic shock
C. increased blood flow
D. complement activation
Answer: C
Critical Thinking Questions
Q. Describe the process of inflammation in an area that has been traumatized, but not infected.
A. The cell debris and damaged cells induce macrophages to begin to clean them up. Macrophages release cytokines that attract neutrophils, followed by more macrophages. Other mediators released by mast cells increase blood flow to the area and also vascular permeability, allowing the recruited cells to get from the blood to the site of infection, where they can phagocytose the dead cells and debris, preparing the site for wound repair.
Q. Describe two early induced responses and what pathogens they affect.
A. Interferons are produced in virally infected cells and cause them to secrete signals for surrounding cells to make antiviral proteins. C-reactive protein is induced to be made by the liver and will opsonize certain species of bacteria.
Glossary
- acute inflammation
- inflammation occurring for a limited time period; rapidly developing
- chemokine
- soluble, long-range, cell-to-cell communication molecule
- chronic inflammation
- inflammation occurring for long periods of time
- complement
- enzymatic cascade of constitutive blood proteins that have antipathogen effects, including the direct killing of bacteria
- cytokine
- soluble, short-range, cell-to-cell communication molecule
- early induced immune response
- includes antimicrobial proteins stimulated during the first several days of an infection
- fas ligand
- molecule expressed on cytotoxic T cells and NK cells that binds to the fas molecule on a target cell and induces it do undergo apoptosis
- granzyme
- apoptosis-inducing substance contained in granules of NK cells and cytotoxic T cells
- histamine
- vasoactive mediator in granules of mast cells and is the primary cause of allergies and anaphylactic shock
- inflammation
- basic innate immune response characterized by heat, redness, pain, and swelling
- interferons
- early induced proteins made in virally infected cells that cause nearby cells to make antiviral proteins
- macrophage
- ameboid phagocyte found in several tissues throughout the body
- mast cell
- cell found in the skin and the lining of body cells that contains cytoplasmic granules with vasoactive mediators such as histamine
- monocyte
- precursor to macrophages and dendritic cells seen in the blood
- neutrophil
- phagocytic white blood cell recruited from the bloodstream to the site of infection via the bloodstream
- opsonization
- enhancement of phagocytosis by the binding of antibody or antimicrobial protein
- pattern recognition receptor (PRR)
- leukocyte receptor that binds to specific cell wall components of different bacterial species
- perforin
- molecule in NK cell and cytotoxic T cell granules that form pores in the membrane of a target cell
- phagocytosis
- movement of material from the outside to the inside of the cells via vesicles made from invaginations of the plasma membrane